A New History of Life (39 page)
Read A New History of Life Online
Authors: Peter Ward

In the Triassic, the crocodile lineage and the dinosaur lineage shared a quadrupedal common ancestor. This beast may have been a reptile from South Africa named
Euparkeria
. This group is technically called the Ornithodira, and from its earliest members this group began to evolve toward bipedalism. This is shown by their anklebones, which simplified into a simple hinge joint from the more complex system found in quadrupeds. This, accompanied by a lengthening of the hind limbs relative to the forelimbs, is also evidence of this, as is the neck, which elongates and forms a slight S shape. These early Ornithodira themselves split into two distinct lineages. One took to the air. These were the pterosaurs, and the late Triassic Ornithodira named
Scleromochlus
might be the very first of its kind; a still-terrestrial form looks like a fast runner that perhaps began gliding between long steps using arms with skin flaps. The oldest undoubted flying pterosaur was
Eudimorphodon
, also of the Late Triassic.
While these ornithodires edge toward flight, their terrestrial sister group headed toward the first dinosaur morphology. The Triassic
Lagosuchus
was a transitional form, being between a bipedal runner and a quadruped. It probably moved slowly on all fours, but reared up on its hind legs for bursts of speed—the bursts necessary to bring down prey, for this was a predator. But it still had forelimbs and hands that had not yet attained the dinosaur type of morphology, so it is not classified as a dinosaur. Its successor, the Triassic
Herrerasaurus
, meets all the requirements and is classified as a dinosaur—the first. But, as we shall see below, it may have lacked one attribute that its immediate descendants would rectify: a new kind of respiratory system that could handle the still-lowering oxygen content of Earth’s atmosphere.
This first dinosaur was fully bipedal,
1
and it could grasp objects with its hands, having a thumb like we do. This five-fingered hand was
distinct from the functionally three-toed foot (there were five actual toes, but two were so vestigial that only three toes touched the ground while running or walking). Because it was not totally bipedal, evolution no longer had to worry about maintaining a hand that had to touch the ground for locomotion. So with a free appendage no longer necessary for locomotion, what to do with it? The much later and more famous
T. rex
reduced the size of the forearm to the point that some have suggested it was nonfunctional. Not so for these first dinosaurs, however. While their posture was that of the later carnivorous dinosaurs so familiar to us, their hands were obviously used—probably for catching and holding prey while on the run.
So this is the body plan of the first dinosaurs, from which all the rest evolved; bipedal, elongated neck, grasping hands with a functional thumb, and a large and distinctive pelvis for the massive muscles and necessary large surface area for these muscles used in walking and running. These early bipeds were relatively small, and before the end of the Triassic they again split into two groups, which remained the most fundamental split of the entire dinosaur clan. A species of these bipedal, Triassic dinosaurs modified its hipbones to incorporate a back-turned pubis, compared to the forward-facing pubis of the first dinosaurs. As any schoolboy knows, this change in pelvic structure marks the division of the dinosaurs into the two great divisions: the ancestral saurischians, and their derived descendants, with whom they would share the world for about the next 170 million years: the ornithischians.
Of interest here, of course, is how dinosaurs breathed.
2
It has been discovered that their respiratory system was quite different from the cold-blooded reptiles we find today, but very similar to that of the warm-blooded birds. The lungs of modern-day amniotes (reptiles, birds, and mammals) are of two basic types (although we will see that there are more than two respiratory systems, which include lungs, circulatory system, and blood pigment type). Both kinds of lungs can be reasonably derived from some single kind of Carboniferous reptilian ancestor that had simple saclike lungs. Extant mammals all have alveolar lungs, while extant turtles, lizards, birds, and crocodiles—all
the rest—have septate lungs. Alveolar lungs consist of millions of highly vascularized, spherical sacs called alveoli. Air flows in and out of the sacs. It is therefore bidirectional.
We mammals use this system, and our familiar breathing—in, out, in, out—is quite typical. Our air has to be pulled into these sacs, and then expelled again as oxygen switches place with carbon dioxide. We do this by a combination of rib cage expansion (powered by muscles, of course) and contraction of the large suite of muscles collectively called the diaphragm. Somewhat paradoxically, contraction of the diaphragm causes the volume of the lungs to increase. These two activities—the interacting rib expansion/diaphragm contraction—create a reduction in air pressure within the lung volume, and air flows in. Exhaling is partially accomplished by elastic rebound of the individual alveoli: when they inflate they enlarge, and soon after they naturally contract due to the elastic properties of their tissue. The many alveoli used in this kind of lung allow for a very efficient oxygen acquisition system, which we warm-blooded mammals very much need in order to maintain our active, movement-rich lifestyles. But the fact that air goes in and out of the same tube is very inefficient and reduces oxygen uptake relative to the amount of energy expended to get it.
3
In contrast to the mammalian lung, the septate lung found in reptiles and birds is like one giant alveolus. To break it into smaller pockets that increases surface area for respiratory exchange, a large number of blade-like sheets of tissue extend into the sac. These partitioning elements are the septa, which give these kinds of lungs their name. There are many variations on this basic lung design among the many different kinds of animals that use it. Some kinds of septate lungs are partitioned into small chambers; others have secondary sacs that rest outside the lung, but are connected to it by tubes. As in the alveolar lung, airflow is bidirectional in most—but, as recently discovered, not all, and the exceptions to this rule, recently found, have profoundly changed our understanding of not only the paleobiology of early reptiles but their fate during the Permian mass extinction.
Septate lungs are not elastic and thus do not naturally contract in size following inhalation. Lung ventilation also varies across groups
with the septate lung. Lizards and snakes use rib movement to draw air in, but as we have seen, locomotion in lizards inhibits complete expansion of the lung cavity, and thus lizards do not breathe while moving.
The variety of modifications of the septate lung makes this system more diverse than the alveolar system. For instance, crocodiles have both a septate lung and a diaphragm—an organ not found in the snakes, lizards, or birds. But the crocodile diaphragm is also somewhat different from that in mammals: it is not muscular, but is attached to the liver, and movement of this liver-diaphragm acts like a piston to inflate the lungs, with muscles attaching to the pelvis. The mammalian (including human) diaphragm pulls the liver in just the same way a crocodilian one does, creating a visceral piston, but the way this is accomplished differs in crocs and mammals.
Until recently, the septate lungs of crocodile and alligator lungs were considered relatively primitive and therefore inefficient. But a radical new finding not only makes us reassess the respiratory ability of the extant forms, but also puts an entirely new view on the reptilian success across the Permian extinction and in the Triassic.
The most inefficient way to breathe is the mammalian way: inhalation and exhalation through the same tube into the lungs. The inefficiency comes from the disorder of the gas molecules as one exhalation finishes and one inhalation starts. In any sort of more rapid breathing, there is a chaotic collision of exhaled air trying to get out before inhalation begins—and quite often the same gas molecules, including volumes of air with more CO
2
and less O
2
, are sucked back in. It has long been thought that the crocs have this problem as well. But a study in 2010 showed that in fact the crocodilians use a separate one-way path that is similar to that in birds and dinosaurs. The revelation is that the ancient Permian- and Triassic-aged stem reptiles, the groups ultimately giving rise to the modern crocodiles and birds, and to the extinct dinosaurs, were also more efficient in their breathing than their therapsid (protomammal) contemporaries. They went through the filter of the Permian extinction with two great competitive advantages; they were cold-blooded, and they could extract more oxygen out of the
air than a mammal or mammal-like reptile. The deck was stacked against us mammals. We never really had a chance in this most consequential competition for not only survivability but for eventual dominance amid the crisis and chaos of mass extinction. The mammals of the Mesozoic eventually would rarely be larger than rats. Probably highly fearful as rats are as well, surrounded by dinosaurs.
The last kind of lung found in terrestrial vertebrates is a variant on the septate lung. The best example of this kind of lung and its associated respiratory system is found in all birds. The lungs themselves in this system are small and somewhat rigid. Thus bird lungs do not greatly expand and contract as ours do on each breath. But the rib cage is very much involved in respiration, and especially those ribs closest to the pelvic region are very mobile in their connection to the bottom of the sternum, and this mobility is quite important in allowing respiration. But these are not the biggest differences. Very much unlike extant reptiles and mammals, these lungs have appendages added to them known as air sacs, and the resultant system of respiration is highly efficient. Here is why. We mammals (and all other nonavians as well) bring air into our dead-end lungs and then exhale it. Birds have a very different system.
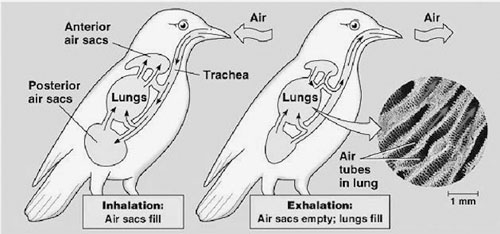
When a bird inspires air, the air goes first into the series of air sacs. It then passes into the lung tissue proper, but in so doing the air passes but one way over the lung, since it is not coming down a trachea but from the attached air sacs. Exhaled air then passes out of the lungs. The one-way flow of air across the lung membranes allows a countercurrent system to be set up: the air passes one direction, and blood in the blood vessels within the lungs passes in the opposite direction. This countercurrent exchange allows for more efficient oxygen extraction and carbon dioxide venting than are possible in dead-end lungs.
Anatomists have been dissecting and describing birds for centuries. It thus seems odd that an accurate understanding of bird air-sac anatomy did not occur until 2005. Two bird anatomists, Patrick O’Connor and Leon Claessens, injected substantial quantities of fast-jelling plastic into the respiratory systems of many different birds and then carefully dissected the corpses and described the anatomy of the filled cavities, the now plastic-filled air sacs.
4
To their surprise, they found that avian air sacs are much more voluminous and complicated than anyone had suspected. For the first time, the real relationship of air sac to bone in pneumatized bone—bones with large cavities in them—could be observed. In the same paper the two authors then compared the anatomy of pneumatized bird bones to pneumatized dinosaur bones. The similarity was remarkable, for there were the same shapes of holes in the same (or homologous) bones.
Those arguing that there was no air-sac system in dinosaurs have not denied that the dinosaur bones had holes in them. They said that the holes were there, all right, but that they were adaptations simply for lightening the bones. But there comes a point when arguing that the similarity is simply coincidence in shape collapses under the weight of too great a coincidence.
In the diagram on page 258, the various air sacs are shown with their communication to the lungs. It is clear that the volume of air sacs far exceeds the volume of the lungs themselves. The air sacs are not involved in removing oxygen; they are an adaptation that allows the countercurrent system to work. There is no question that the greater
efficiency of this system compared to all other lungs in vertebrates is related to the two-cycle, countercurrent system produced by the air-sac lung anatomy in birds.
By 2005, the evidence that many dinosaurs had air sacs was overwhelming. Until then, one group of anatomists had vigorously argued that dinosaur lungs were no different than modern crocodile lungs, just large, and that the avian lung,
5
with its many auxiliary air sacs as well as a one-way airflow, did not appear until the Cretaceous, some 100 million years ago—and then was found only in birds! That view no longer became tenable. But in 2005 there was still no appreciation of the degree to which atmospheric oxygen levels had changed in the Early Mesozoic, or that such changes might have had any influence at all on the evolution of these various respiratory systems.