Janus (31 page)
Authors: Arthur Koestler

,
we can hear a note of sharp dissent from the orthodox attitude:
The Origin
of Species
this introduction no longer appears.
Chapter I, 10
, I mentioned the classic example of
the forelimbs of vertebrates which, whether they serve reptiles, birds,
whales or man, show the same basic design of bones, muscles, nerves, etc.,
and are accordingly called homologous organs. The functions of legs,
wings and flippers are quite different, yet they all are variations on
a single theme -- strategic modifications of a pre-existing structure:
the forelimb of the common reptilian ancestor. Once Nature has 'taken
out a patent' on a vital organ, she sticks to it, and that organ becomes
a stable evolutionary holon. Its basic design seems to be governed by
a fixed
evolutionary canon
; while its adaptation to swimming,
walking, or flying is a matter of evolution's flexible
strategy
.
hierarchy, from the sub-cellular level to the primate brain. The same four
chemical bases in the chromosomal nucleic acid -- DNA -- constitute the
four-letter alphabet of the genetic codes throughout the animal kingdom;
the same 'make' of organelles function in their cells; the same chemical
fuel -- ATP -- provides their energy; the same contractile proteins serve
the motions of the amoeba and of human muscles. Animals and plants are
made of homologous molecules, organelles, and even more complex homologous
sub-structures. They are the stable holons in the evolutionary flux,
the nodes on the tree of life.
concerned with the nature of evolutionary
strategies
(Darwinian,
Lamarckian, etc.) which made the higher forms of life branch out of
the roots at the base of the hierarchy. But dazzled by the prodigious
variety
of plants and animals, biologists were inclined to pay less
attention to the
uniformity
of those basic units -- reflected in
the phenomena of homology -- and the
limitations
which it imposed
on all existing and possible forms of life on this planet. After all,
the basic uniformity of the organelles which constitute the living cell
is itself derived from the limitations imposed by the basic chemistry
of organic matter such as amino-acids, proteins, enzymes. On a higher
level,' the genetic micro-hierarchies impose further constraints on
hereditary variations. Still further up the 'great central something'
regulates -- in ways unknown to us -- the 'harmonious coordination' of
genetic changes. Their combined effect is the evolutionary canon, which
permits a great amount of variations, but only in limited directions
on
a limited number of themes
. Evolution is not a free-for-all but -- to
revert to our formula -- a game with fixed rules and flexible strategies,
played over thousands of millennia.
use the example of the Australian marsupials, which I used in
The Ghost
in the Machine
.* I called them an enigma wrapped in a puzzle. The enigma
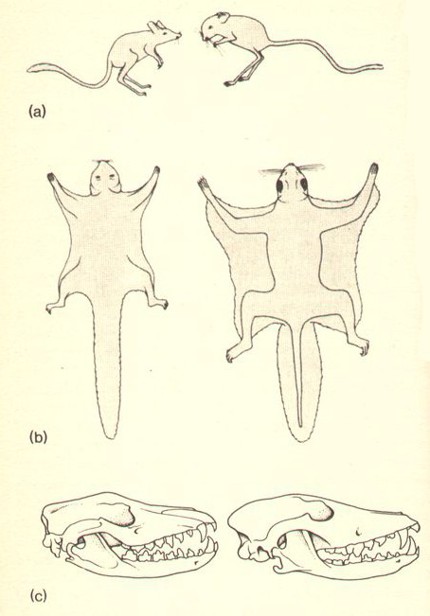
is shown by the drawings on p. 208. The puzzle is why evolutionists
refuse to see the problems that it poses.
They have evolved, independently from each other, from a common ancestry
(the now extinct therapsids, or mammal-like reptiles). The marsupial
embryo is expelled from the womb in a very immature state of development
and is reared in an elastic pouch attached to the mother's belly. A newborn
kangaroo is a half-finished job: about an inch long, naked, blind,
its hind-legs no more than embryonic buds. One might speculate whether
the human infant, more developed but still helpless at birth, would
be better off in a maternal pouch; one is also reminded of African
or Japanese women carrying their infants strapped to their backs. But
whether the marsupial method is better or worse than the placental, the
point is that they differ. Pouch and placenta might be called variations
in strategy within the general schema of mammalian reproduction.
evolution some time before Australia became separated from the Asiatic
mainland in the late Cretacean. The marsupials (who had branched out from
the common ancestral type earlier than the placentals) got into Australia
before it was cut off; the placentals did not. So the two lines evolved
in complete separation for about a hundred million years. The enigma is
why so many animals in the Australian fauna, produced by the independent
evolutionary line of the marsupials, look so startlingly like their
opposite numbers among placentals. The drawings on p. 208 show on the
left side three specimens of marsupials, on the right the corresponding
placentals. It is as if two artists who have never met and never shared
the same model, had drawn parallel series of almost identical portraits.
managed to get there in time were tiny, mouse-like, pouched animals,
perhaps not unlike the still extant yellow-footed pouched mouse,
but even more primitive. And yet these archaic creatures, confined to
their island continent, branched out and gave rise to pouched versions
of our placental moles, ant-eaters, flying squirrels, cats, wolves,
lions, and so on -- each like a somewhat clumsy copy of its placental
namesake. Why, if evolution were a free-for-all, why did Australia not
produce some entirely different species of animals, like the bug-eyed
monsters of science fiction? The only moderately unorthodox creation of
that isolated island in a hundred million years are the kangaroos and
wallabies; the rest of the fauna consists of rather inferior duplicates
of more efficient placental types -- variations on a limited number of
themes, within the repertory of the evolutionary canon.
offer is summed up in the following quotation from an authoritative
textbook:
the same problem, concludes that the explanation is 'selection of random
mutations'. [2]
that the vague phrase 'preying on animals of approximately the same size
and habits' -- which can be applied to hundreds of different species
-- provides a sufficient explanation for the emergence of the nearly
identical skulls shown on p. 208. Even the evolution of a single species
of wolf by random mutation plus selection presents, as we have seen,
insurmountable difficulties. To
duplicate
this process independently
on island and mainland would mean squaring a miracle. The puzzle remains
why the Darwinians are not puzzled -- or pretend not to be.*
Doppelgängers
lend strong support to
the hypothesis that there are unitary laws underlying evolutionary
diversity, which permit virtually unlimited variations on a limited
number of themes. They include, on the lower levels of the hierarchy,
macromolecules, organelles and cells which represent evolutionary holons;
higher up, homologous organs such as the vertebrate forelimbs, lungs, and
gills, not to mention eyes equipped with lenses -- which have evolved,
independently from each other, several times in evolutionary lines
as far apart as molluscs, spiders and vertebrates. Still higher up we
have to include in the list the more or less standardized vertebrate
types exemplified by the drawings. The 'more or less' we can ascribe to
variations in evolutionary strategy in a changing environment; but their
standardization we can only explain by rules built into the genetic
micro-hierarchies which confine evolutionary advances to certain main
avenues, and filter out the rest.
transcendentalists of the eighteenth century, including Goethe
(and eventually to Plato); but it was revived by a number of modern
evolutionists who toyed with the idea of 'internal selection' without
spelling out its profound implications.* Thus Helen Spurway concluded from
the universal recurrence of homologous forms that the organism has only
'a restricted mutation spectrum' which 'determines its possibilities
of evolution'.
[3]
Other biologists have talked of 'organic
laws co-determining evolution', 'moulding influences guiding evolutionary
change along certain avenues'
[4]
; while Waddington reverted
to 'the notion of archetypes . . . the idea, that is, that there are
only a certain number of basic patterns which organic form can assume'.
[5]
What they are implying (without saying it in as many
words) is that, given the conditions on our particular planet, its
gravity and temperature; the composition of its atmosphere, oceans and
soil; the nature of available energies and raw materials, life from its
inception in the first blob of living slime could only evolve
in a
limited number of directions in a limited number of ways
. But this
in turn implies that just as the basic pattern of the twin wolves was
foreshadowed, or present
in potentia
, in their common ancestry,
so the mammal-like reptile creature must have been potentially present
in the ancestral chordate -- and so on back to the ancestral protist,
and the first self-replicating strand of nucleic acid.
homology -- which Sir Alister Hardy has called 'absolutely fundamental
to what we are talking about when we speak of evolution'.
[6]
If this line of argument is correct, it puts an end to the monsters
of science fiction as possible forms of life on earth -- or on other
planets similar to it. But it does not mean the opposite either:
it emphatically does not mean a rigidly predetermined universe which
unwinds like a mechanical clockwork. It means -- to revert to one of
the leitmotifs of this book -- that the evolution of life is a splendid
game played according to fixed rules which limit its possibilities but
leave sufficient scope for virtually limitless variations. The rules
are inherent in the basic structure of living matter, the variations are
derived from flexible strategies which take advantage of the opportunities
offered by the former.
alone, nor the execution of a rigidly predetermined computer programme.
It could be compared to musical composition of the classical type, whose
possibilities are limited by the rules of harmony and the structure of
the diatonic scales -- which nevertheless permit an inexhaustible number
of original creations. Or it could be compared to a game of chess,
obeying fixed rules with equally inexhaustible variations. And lastly --
to quote from
we can hear a note of sharp dissent from the orthodox attitude:
This situation, where scientific men rally to the defence of a doctrine
they are unable to define scientifically, much less demonstrate with
scientific rigour, attempting to maintain its credit with the public
by the suppression of criticism and the elimination of difficulties,
is abnormal and undesirable in science. [18]
The Origin
of Species
this introduction no longer appears.
Chapter I, 10
, I mentioned the classic example of
the forelimbs of vertebrates which, whether they serve reptiles, birds,
whales or man, show the same basic design of bones, muscles, nerves, etc.,
and are accordingly called homologous organs. The functions of legs,
wings and flippers are quite different, yet they all are variations on
a single theme -- strategic modifications of a pre-existing structure:
the forelimb of the common reptilian ancestor. Once Nature has 'taken
out a patent' on a vital organ, she sticks to it, and that organ becomes
a stable evolutionary holon. Its basic design seems to be governed by
a fixed
evolutionary canon
; while its adaptation to swimming,
walking, or flying is a matter of evolution's flexible
strategy
.
hierarchy, from the sub-cellular level to the primate brain. The same four
chemical bases in the chromosomal nucleic acid -- DNA -- constitute the
four-letter alphabet of the genetic codes throughout the animal kingdom;
the same 'make' of organelles function in their cells; the same chemical
fuel -- ATP -- provides their energy; the same contractile proteins serve
the motions of the amoeba and of human muscles. Animals and plants are
made of homologous molecules, organelles, and even more complex homologous
sub-structures. They are the stable holons in the evolutionary flux,
the nodes on the tree of life.
concerned with the nature of evolutionary
strategies
(Darwinian,
Lamarckian, etc.) which made the higher forms of life branch out of
the roots at the base of the hierarchy. But dazzled by the prodigious
variety
of plants and animals, biologists were inclined to pay less
attention to the
uniformity
of those basic units -- reflected in
the phenomena of homology -- and the
limitations
which it imposed
on all existing and possible forms of life on this planet. After all,
the basic uniformity of the organelles which constitute the living cell
is itself derived from the limitations imposed by the basic chemistry
of organic matter such as amino-acids, proteins, enzymes. On a higher
level,' the genetic micro-hierarchies impose further constraints on
hereditary variations. Still further up the 'great central something'
regulates -- in ways unknown to us -- the 'harmonious coordination' of
genetic changes. Their combined effect is the evolutionary canon, which
permits a great amount of variations, but only in limited directions
on
a limited number of themes
. Evolution is not a free-for-all but -- to
revert to our formula -- a game with fixed rules and flexible strategies,
played over thousands of millennia.
use the example of the Australian marsupials, which I used in
The Ghost
in the Machine
.* I called them an enigma wrapped in a puzzle. The enigma
is shown by the drawings on p. 208. The puzzle is why evolutionists
refuse to see the problems that it poses.
* The section that follows is a compressed version of The Ghost in
the Machine, pp. 143-6.
(a) Marsupial jerboa and placental jerboa
(b) Marsupial flying phalanger and placental flying squirrel.
(c) Skull of Tasmanian wolf and skull of placental wolf (after Hardy).
They have evolved, independently from each other, from a common ancestry
(the now extinct therapsids, or mammal-like reptiles). The marsupial
embryo is expelled from the womb in a very immature state of development
and is reared in an elastic pouch attached to the mother's belly. A newborn
kangaroo is a half-finished job: about an inch long, naked, blind,
its hind-legs no more than embryonic buds. One might speculate whether
the human infant, more developed but still helpless at birth, would
be better off in a maternal pouch; one is also reminded of African
or Japanese women carrying their infants strapped to their backs. But
whether the marsupial method is better or worse than the placental, the
point is that they differ. Pouch and placenta might be called variations
in strategy within the general schema of mammalian reproduction.
* Not counting the nearly extinct egg-laying mammals, such as the
duck-billed platypus.
evolution some time before Australia became separated from the Asiatic
mainland in the late Cretacean. The marsupials (who had branched out from
the common ancestral type earlier than the placentals) got into Australia
before it was cut off; the placentals did not. So the two lines evolved
in complete separation for about a hundred million years. The enigma is
why so many animals in the Australian fauna, produced by the independent
evolutionary line of the marsupials, look so startlingly like their
opposite numbers among placentals. The drawings on p. 208 show on the
left side three specimens of marsupials, on the right the corresponding
placentals. It is as if two artists who have never met and never shared
the same model, had drawn parallel series of almost identical portraits.
managed to get there in time were tiny, mouse-like, pouched animals,
perhaps not unlike the still extant yellow-footed pouched mouse,
but even more primitive. And yet these archaic creatures, confined to
their island continent, branched out and gave rise to pouched versions
of our placental moles, ant-eaters, flying squirrels, cats, wolves,
lions, and so on -- each like a somewhat clumsy copy of its placental
namesake. Why, if evolution were a free-for-all, why did Australia not
produce some entirely different species of animals, like the bug-eyed
monsters of science fiction? The only moderately unorthodox creation of
that isolated island in a hundred million years are the kangaroos and
wallabies; the rest of the fauna consists of rather inferior duplicates
of more efficient placental types -- variations on a limited number of
themes, within the repertory of the evolutionary canon.
offer is summed up in the following quotation from an authoritative
textbook:
Tasmanian [i.e., marsupial] and true wolves are both running predators,
preying on other animals of about the same size and habits. Adaptive
similarity [i.e., adaptation to similar environments] involves
similarity also of structure and function. The mechanism of such
evolution is natural selection. [1]
the same problem, concludes that the explanation is 'selection of random
mutations'. [2]
that the vague phrase 'preying on animals of approximately the same size
and habits' -- which can be applied to hundreds of different species
-- provides a sufficient explanation for the emergence of the nearly
identical skulls shown on p. 208. Even the evolution of a single species
of wolf by random mutation plus selection presents, as we have seen,
insurmountable difficulties. To
duplicate
this process independently
on island and mainland would mean squaring a miracle. The puzzle remains
why the Darwinians are not puzzled -- or pretend not to be.*
* Various terms have been invented to describe this phenomenon such as
'convergence', 'parallelism', 'homeoplasy', but these are purely
descriptive, without explanatory value.
Doppelgängers
lend strong support to
the hypothesis that there are unitary laws underlying evolutionary
diversity, which permit virtually unlimited variations on a limited
number of themes. They include, on the lower levels of the hierarchy,
macromolecules, organelles and cells which represent evolutionary holons;
higher up, homologous organs such as the vertebrate forelimbs, lungs, and
gills, not to mention eyes equipped with lenses -- which have evolved,
independently from each other, several times in evolutionary lines
as far apart as molluscs, spiders and vertebrates. Still higher up we
have to include in the list the more or less standardized vertebrate
types exemplified by the drawings. The 'more or less' we can ascribe to
variations in evolutionary strategy in a changing environment; but their
standardization we can only explain by rules built into the genetic
micro-hierarchies which confine evolutionary advances to certain main
avenues, and filter out the rest.
transcendentalists of the eighteenth century, including Goethe
(and eventually to Plato); but it was revived by a number of modern
evolutionists who toyed with the idea of 'internal selection' without
spelling out its profound implications.* Thus Helen Spurway concluded from
the universal recurrence of homologous forms that the organism has only
'a restricted mutation spectrum' which 'determines its possibilities
of evolution'.
[3]
Other biologists have talked of 'organic
laws co-determining evolution', 'moulding influences guiding evolutionary
change along certain avenues'
[4]
; while Waddington reverted
to 'the notion of archetypes . . . the idea, that is, that there are
only a certain number of basic patterns which organic form can assume'.
[5]
What they are implying (without saying it in as many
words) is that, given the conditions on our particular planet, its
gravity and temperature; the composition of its atmosphere, oceans and
soil; the nature of available energies and raw materials, life from its
inception in the first blob of living slime could only evolve
in a
limited number of directions in a limited number of ways
. But this
in turn implies that just as the basic pattern of the twin wolves was
foreshadowed, or present
in potentia
, in their common ancestry,
so the mammal-like reptile creature must have been potentially present
in the ancestral chordate -- and so on back to the ancestral protist,
and the first self-replicating strand of nucleic acid.
* See above, Ch. IX, 7. For an excellent short
critical discussion see L. L. Whyte's Internal Factors in
Evolution and W. H. Thorpe's review of the book in Nature,
14 May, 1966.
homology -- which Sir Alister Hardy has called 'absolutely fundamental
to what we are talking about when we speak of evolution'.
[6]
If this line of argument is correct, it puts an end to the monsters
of science fiction as possible forms of life on earth -- or on other
planets similar to it. But it does not mean the opposite either:
it emphatically does not mean a rigidly predetermined universe which
unwinds like a mechanical clockwork. It means -- to revert to one of
the leitmotifs of this book -- that the evolution of life is a splendid
game played according to fixed rules which limit its possibilities but
leave sufficient scope for virtually limitless variations. The rules
are inherent in the basic structure of living matter, the variations are
derived from flexible strategies which take advantage of the opportunities
offered by the former.
alone, nor the execution of a rigidly predetermined computer programme.
It could be compared to musical composition of the classical type, whose
possibilities are limited by the rules of harmony and the structure of
the diatonic scales -- which nevertheless permit an inexhaustible number
of original creations. Or it could be compared to a game of chess,
obeying fixed rules with equally inexhaustible variations. And lastly --
to quote from
Other books
Repairman Jack [07]-Gateways by F. Paul Wilson
Under Cover of Daylight by James W. Hall
Listed: Volume III by Noelle Adams
Steam (Legends Saga Book 3) by Stacey Rourke
Miss Cresswell's London Triumph by Evelyn Richardson
NaGeira by Paul Butler
The First 90 Days by Michael Watkins
Fall of a Philanderer by Carola Dunn
Head Full of Mountains by Brent Hayward