The Origin of Humankind (4 page)
Read The Origin of Humankind Online
Authors: Richard Leakey

The valley has dramatic highlands with cool, forested plateaus, and precipitous slopes that plunge 3000 feet to hot, arid lowlands. Biologists have come to realize that mosaic environments of this kind, which offer many different kinds of habitat, drive evolutionary innovation. Populations of a species that once were widespread and continuous may become isolated and exposed to new forces of natural selection. Such is the recipe for evolutionary change. Sometimes that change is toward oblivion, if favorable environments disappear. This, clearly, was the fate of most of the African apes: just three species exist today—the gorilla, the common chimpanzee, and the pygmy chimpanzee. But while most ape species suffered because of the environmental shift, one of them was blessed with a new adaptation that allowed it to survive and prosper. This was the first bipedal ape. Being bipedal clearly bestowed important survival advantages in the changing conditions. The job of anthropologists is to discover what those advantages were.
Anthropologists tend to view the importance of bipedality in human evolution in two ways: one school emphasizes the freeing up of the forelimbs for carrying things; the other emphasizes the fact that bipedalism is a more energy efficient mode of locomotion, and sees the ability to carry things merely as a fortuitous by-product of the upright stance.
The first of these two hypotheses was proposed by Owen Lovejoy and published in a major paper in
Science
in 1981. Bipedalism, he argued, is an inefficient mode of locomotion, so it must have evolved for carrying things. How could the ability to carry things give bipedal apes a competitive edge over other apes?
Ultimately, evolutionary success depends on producing surviving offspring, and the answer, suggested Lovejoy, lay in the opportunity that this new ability gave male apes to boost the reproductive rate of the female, by gathering food for her. Apes, he pointed out, reproduce slowly, having one infant every four years. If human females had access to more energy—that is, food—they might successfully produce more offspring. If a male helped provide a female with more energy by collecting food for her and for her offspring, she would be able to boost her reproductive output.
There would be a further biological consequence of the male’s activity, this time in the social realm. Since it would not benefit the male in a Darwinian sense to provision a female unless he were sure she was producing his offspring, Lovejoy suggested that the first human species was monogamous, with the nuclear family emerging as a way of increasing reproductive success, and thus outcompeting the other apes. He supported his argument by further biological analogy. In most primate species, for example, males compete with each other for sexual control of as many females as possible. They often fight with one another during this process, and are endowed with large canine teeth, which they use as weapons. Gibbons are rare in that they form male-female pairs, and—presumably because they do not have reason to fight with one another—the males have small canine teeth. The small canines in the earliest humans may be an indication that, like gibbons, they formed male-female pairs, Lovejoy argued. The social and economic bonds of the provisioning arrangement would in turn have driven an increase in the size of the brain.
Lovejoy’s hypothesis, which enjoyed considerable attention and support, is powerful because it appeals to fundamental biological issues, not cultural ones. It has weak points, however; for one thing, monogamy is not a common social arrangement among technologically primitive people. (Only 20 percent of such societies are monogamous.) The hypothesis was therefore criticized for seeming to draw on a trait of Western society, not one of hunter-gatherers. A second criticism, perhaps more serious, is that the males of the known early human species were about twice the size of females. In all species of primate that have been studied, this great difference in body size, known as dimorphism, correlates with polygyny, or competition among the males for access to females; dimorphism is not seen in monogamous species. For me, this fact alone is sufficient to sink a promising theoretical approach, and an explanation other than monogamy must be sought for the small canines. One possibility is that the mechanism of masticating food required a grinding rather than a slicing motion; large canines would impair such a motion. Lovejoy’s hypothesis enjoys less support now than it did a decade ago.
The second major bipedalism theory is much more persuasive, partly for its simplicity. Proposed by the anthropologists Peter Rodman and Henry McHenry, of the University of California, Davis, the hypothesis states that bipedalism was advantageous in the changing environmental conditions because it offered a more efficient means of locomotion. As the forests dwindled, food resources in woodland habitats, such as fruit trees, would have become too dispersed to be efficiently exploitable by conventional apes. According to this hypothesis, the first bipedal apes were human only in their mode of locomotion. Their hands, jaws, and teeth would have remained apelike, because their diet had not changed, only their manner of procuring it.
To many biologists, this proposal initially seemed unlikely; researchers at Harvard University had shown some years earlier that walking on two legs is less efficient than walking on four. (This shouldn’t be a surprise to anyone with a dog or a cat; both run, embarrassingly, much faster than their owners.) The Harvard researchers had, however, compared the energy efficiency of bipedalism in humans with that of quadrupedalism in horses and dogs. Rodman and McHenry pointed out that the proper comparison should have been between humans and chimpanzees. When these comparisons are done, it turns out that bipedalism in humans is more efficient than quadrupedalism in chimpanzees. An energy-efficiency argument as a force of natural selection in favor of bipedalism, they concluded, is therefore plausible.
There have been many other suggestions for the factors that drove the evolution of bipedalism, such as the need to look over tall grass while monitoring predators and the need to adopt a more efficient posture for cooling during daytime foraging. Of them all, I find Rodman and McHenry’s the most cogent, because it is firmly biologically based and fits the ecological changes that were occurring when the first human species evolved. If the hypothesis is correct, it will mean that when we find fossils of the first human species, we may fail to recognize them as such, depending on which bones we have. If the bones are those of the pelvis or lower limbs, then the bipedal mode of locomotion will be evident, and we will be able to say “human.” But if we were to find certain parts of the skull, jaw, or some teeth, they might look just like those of an ape. How would we know whether they were those of a bipedal ape or a conventional ape? It’s an exciting challenge.
If we could visit the Africa of 7 million years ago to observe the behavior of the first humans, we would see a pattern more familiar to primatologists, who study the behavior of monkeys and apes, than to anthropologists, who study the behavior of humans. Rather than living as aggregations of families in nomadic bands, as modern hunter-gatherers do, the first humans probably lived like savanna baboons. Troops of thirty or so individuals would forage in a coordinated way over a large territory, returning to favored sleeping places at night, such as cliffs or clumps of trees. Mature females and their offspring would make up most of the troop’s numbers, with just a few mature males present. The males would be continually looking for mating opportunities, with the dominant individuals achieving the most success. Immature and low-ranking males would be very much on the periphery of the troop, often foraging by themselves. The individuals in the troop would have the human aspect of walking bipedally but would be behaving like savanna primates. Ahead of them lay 7 million years of evolution—a pattern of evolution that was complex, as we shall see, and by no means certain. For natural selection operates according to immediate circumstances and not toward a long-term goal.
Homo sapiens
did eventually evolve as a descendant of the first humans, but there was nothing inevitable about it.
A CROWDED FAMILY
B
y my count, fossil specimens of varying degrees of incompleteness, representing at least a thousand individuals of various human species, have been recovered from South and East Africa from the earliest part of the record—that is, from about 4 million years ago up until almost a million years ago (many more have been found in the later record). The oldest human fossils found in Eurasia may be close to 2 million years old. (The New World and Australia were populated much more recently, some 20,000 and 55,000 years ago, respectively.) It is fair to say, therefore, that most of the action of human prehistory took place in Africa. The questions anthropologists must answer about this action are twofold: First, what species populated the human family tree between 7 million years ago and 2 million years ago, and how did they live? Second, how were the species related to each other evolutionarily? In other words, what was the shape of the family tree?
My anthropological colleagues face two practical challenges in addressing these problems. The first is what Darwin called “the extreme imperfection of the geological record.” In his
Origin of Species
, Darwin devoted an entire chapter to the frustrating gaps in the record, which result from the capricious forces of fossilization and later exposure of bones. The conditions that favor the rapid burial and possible fossilization of bones are rare. And ancient sediments may become uncovered through erosion—when a stream cuts through them, for instance—but which pages of prehistory are reopened in this way is purely a matter of chance, and many of the pages remain hidden from view. For instance, in East Africa, the most promising repository for early human fossils, there are very few fossil-bearing sediments from the period between 4 million and 8 million years ago. This is a crucial period in human prehistory, because it includes the origin of the human family. Even for the time period after 4 million years we have far fewer fossils than we would like.
The second challenge stems from the fact that the majority of fossil specimens discovered are small fragments—a piece of cranium, a cheekbone, part of an arm bone, and many teeth. The identification of species from meager evidence of this nature is no easy task and is sometimes impossible. The resulting uncertainty allows for many differences of scientific opinion, both in identifying species and in discerning the interrelatedness of species. This area of anthropology, known as taxonomy and systematics, is one of the most contentious. I will avoid the details of the many debates and concentrate instead on describing the overall shape of the tree.
Knowledge of the human fossil record in Africa developed slowly, beginning in 1924 when Raymond Dart announced the discovery of the famous Taung child. Comprising the incomplete skull of a child—part of the cranium, face, lower jaw, and brain case—the specimen was so named because it was recovered from the Taung limestone quarry, in South Africa. Although no precise dating of the quarry sediments was possible, scientific estimates suggest that the child lived about 2 million years ago.
While the Taung child’s head had many apelike features, such as a small brain and a protruding jaw, Dart recognized human aspects too: the jaw protruded less than it does in apes, the cheek teeth were flat, and the canine teeth were small. A key piece of evidence was the position of the foramen magnum—the opening at the base of the skull through which the spinal cord passes into the spinal column. In apes, the opening is relatively far back in the base of the cranium, while in humans it is much closer to the center; the difference reflects the bipedal posture of humans, in which the head is balanced atop the spine, in contrast to ape posture, in which the head leans forward. The Taung child’s foramen magnum was in the center, indicating that the child was a bipedal ape.
Although Dart was convinced of the hominid status of the Taung child, almost a quarter of a century was to pass before professional anthropologists accepted the fossil individual as a human ancestor and not just an ancient ape. The prejudice against Africa as the site of human evolution and a general revulsion at the idea that anything so apelike might be a part of human ancestry combined to consign Dart and his discovery to anthropological oblivion for a long time. By the time anthropologists recognized their error—in the late 1940s—Dart had been joined by the Scotsman Robert Broom, and the two men had found scores of early human fossils from four cave sites in South Africa: Sterkfontein, Swartkrans, Kromdraai, and Maka-pansgat. Following the anthropological custom of the time, Dart and Broom applied a new species name to virtually every fossil they discovered, so that very soon it appeared that there had been a veritable zoo of human species living in South Africa between 3 million and 1 million years ago.
By the 1950s, anthropologists decided to rationalize the plethora of proposed hominid species and recognized just two. Both were bipedal apes, of course, and both were apelike in the way that the Taung child was. The principal difference between the two species was in their jaws and teeth: in both, these were large, but one of the creatures was a more massive version of the other. The more gracile species was given the name
Australopithecus africanus
, which was the appellation Dart had given to the Taung child in 1924; the term means “southern ape from Africa.” The more robust species was called, appropriately,
Australopithecus robustus
(see
figure 2.1
).
From the structure of their teeth, it was obvious that both
africanus
and
robustus
had lived mostly on plant foods. Their cheek teeth were not those of apes—which have pointed cusps, suited to a diet of relatively soft fruit and other vegetation—but were flattened into grinding surfaces. If, as I suspect, the first human species had lived on apelike diets, they would have had apelike teeth. Clearly, by 2 million to 3 million years ago the human diet had changed to one of tougher foods, such as hard fruits and nuts. Almost certainly this indicated that the australopithecines lived in a drier environment than that of apes. The huge size of the robust species’ molars suggests that the food it ate was especially tough and needed extensive grinding; not for nothing are they referred to as “millstone molars.”
The first early human fossil in East Africa was found by Mary Leakey, in August 1959. After almost three decades of searching the sediments of Olduvai Gorge, she was rewarded with the sight of millstone molars, like those of the robust australopithecine species in South Africa. The Olduvai individual was, however, even more robust than its South African cousin. Louis Leakey, who, with Mary, had taken part in the long search, named it
Zinjanthropus boisei:
the generic name means “East African man” and
boisei
referred to Charles Boise, who supported my father and mother in their work at Olduvai Gorge and elsewhere. In the first application of modern geological dating to anthropology, Zinj, as the individual became known, was determined to have lived 1.75 million years ago. Zinj’s name was eventually changed to
Australopithecus boisei
, on the assumption that it was an East African version, or geographical variant, of
Australopithecus robustus
.
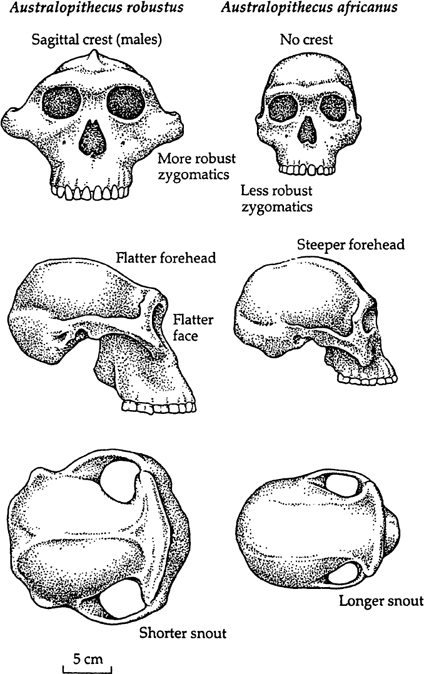
Australopithecine cousins. The principal difference between
Australopithecus robustus
(and
boisei)
and
africanus
is in the chewing mechanisms, which include the structure of the jaws, cheekbones, and associated sites for muscle attachments. The
robustus
species was adapted to a diet that contained tough plant foods, requiring heavy mastication. (Courtesy of A. Walker and R. E. F.
Leakey/Scientific American
, 1978, all rights reserved.)
The names are not particularly important in themselves. What is important is that we are seeing several human species with the same fundamental adaptation, that of bipedalism, a small brain, and relatively large cheek teeth. This was what I saw in the cranium I found resting on a dry streambed on my first expedition to the eastern shore of Lake Turkana, in 1969.
We know from the size of various bones of the skeleton that the males of the australopithecine species were much bigger than the females. They stood more than 5 feet tall, while their mates barely achieved 4 feet. The males must have weighed almost twice as much as the females, a difference of the sort that we see today in some species of savanna baboons. It’s a fair guess, therefore, that the social organization of australopithecines was similar to that of baboons, with dominant males competing for access to mature females, as noted in the previous chapter.
The story of human prehistory became a little more complicated a year after the discovery of Zinj, when my older brother, Jonathan, found a piece of the cranium of another type of hominid, again at Olduvai Gorge. The relative thinness of the cranium indicated that this individual was of slighter build than any of the known australopithecine species. It had smaller cheek teeth and, most significant of all, its brain was almost 50 percent larger. My father concluded that although the australopithecines were part of human ancestry, this new specimen represented the lineage that eventually gave rise to modern humans. Amid an uproar of objections from his professional colleagues, he decided to name it
Homo habilis
, making it the first early member of the genus to be identified. (The name
Homo habilis
, which means “handy man,” was suggested to him by Raymond Dart, and it refers to the supposition that the species were toolmakers.)
The uproar was based on esoteric considerations in many ways; it erupted in part because in order to assign the appellation
Homo
to the new fossil, Louis had to modify the accepted definition of the genus. Until that time, the standard definition, proposed by the British anthropologist Sir Arthur Keith, stated that the brain capacity of the genus
Homo
should equal or exceed 750 cubic centimeters, a figure that was intermediate between that of modern humans and apes; it had become known as the cerebral Rubicon. Despite the fact that the newly discovered fossil from Olduvai Gorge had a brain capacity of only 650 cubic centimeters, Louis judged it to be
Homo
because of its more humanlike (that is, less robust) cranium. He therefore proposed shifting the cerebral Rubicon to 600 cubic centimeters, thereby admitting the new Olduvai hominid to the genus
Homo
. This tactic surely raised the emotional level of the vigorous debate that ensued. Ultimately, however, the new definition was accepted. (It later developed that 650 cubic centimeters is rather small for the average adult brain size in
Homo habilis;
800 cubic centimeters is a closer figure.)
Scientific names aside, the important point here is that the pattern of evolution beginning to emerge from these findings was of two basic types of early human. One type had a small brain and large cheek teeth (the various australopithecine species); the second type had an enlarged brain and smaller cheek teeth
(Homo)
(see
figure 2.2
). Both types were bipedal apes, but something extraordinary had clearly happened in the evolution of
Homo
. We will explore this “something” more fully in the next chapter. In any case, anthropologists’ understanding of the shape of the family tree at this point in human history—that is, at around 2 million years ago—was rather simple.
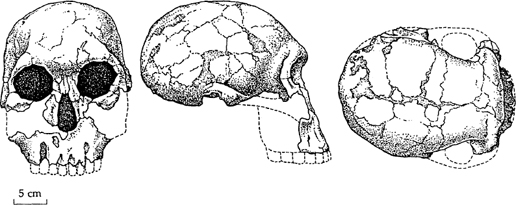
Early
Homo
. This fossil, known by its museum acquisition number of 1470, was found in Kenya in 1972. It lived almost 2 million years ago and is the most complete early specimen of
Homo habilis;
it shows significant brain expansion and reduction in tooth size, compared with the australopithecines. (Courtesy of A. Walker and R. E. F. Leakey
/Scientific American
, 1978, all rights reserved.)