What Mad Pursuit (16 page)
Authors: Francis Crick

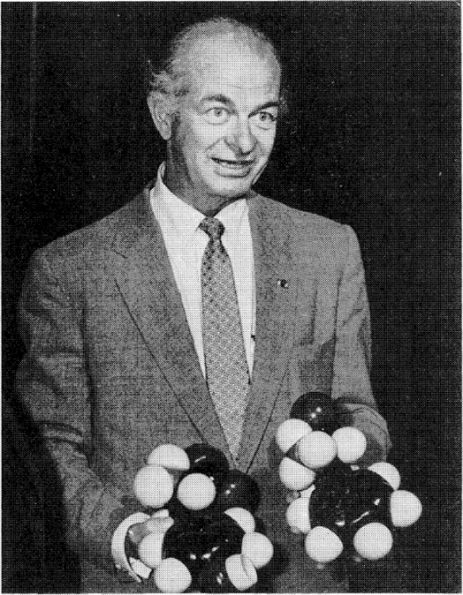
Linus Pauling in the 1950s, holding models of two molecules.
(Courtesy of the Archives, California Institute of Technology.)

Max Delbrück in conversation, June 1959.
(Courtesy of the Archives,
California Institute of Technology.)

Meeting of the RNA Tie Club
[from left to right:
myself, Alex Rich,
Leslie Orgel, Jim Watson).
(Courtesy of Alex Rich, Cambridge, Mass.)

Max Perutz (
right
) handing over
the MRC Laboratory of Molecular Biology
to Sydney Brenner in 1979.
(Author’s collection.)

Maurice Wilkins, Max Perutz,
myself, John Steinbeck, Jim Watson, and John Kendrew
at the Nobel ceremony, 1962.
(Courtesy of Svenskt
Pressfoto, Stockholm.)

King Gustaf VI Adolf of Sweden, with Odile at the Nobel banquet, 1962.
(Courtesy of Svenskt Pressfoto, Stockholm.)

Myself dancing with my elder daughter Gabrielle
at the Nobel Prize celebration, 1962.
(Courtesy of International Magazine
Service, Stockholm.)
The example just given about DNA was a hypothetical one, but I have been caught in this way more than once. Experiments had shown that transfer RNA (tRNA) molecules existed, that amino acids were associated with them, and that there were probably many types of tRNA molecules, each with its own particular amino acid. The obvious next step was to purify at least one type of tRNA away from all the others so that more could be learned about it, as it was obviously better to work where possible on a pure species than on a mixture.
The problem was how to fractionate such a mixture. I argued to myself that since all tRNA molecules had to do a similar job and in particular to fit into the same place, or set of places, at the ribosome, they would all be very similar to each other and thus difficult to separate. The only way to separate them, I felt, was to use some method that tried to latch onto the amino acid joined to the RNA, by going for the particular side group of that amino acid and choosing one, such as cysteine, that was chemically both active and unique. I even tried to do this experimentally.
The argument was not totally silly, but it turned out I was wrong. Though I could not know it at the time, most tRNA molecules have many modified bases. These modifications alter their chromatographic behavior and so make it possible to separate them by much simpler fractionation methods since, in the first instance, only one of them is wanted. There is no need to specify in advance
which
tRNA to study, one simply experiments on the one that is easiest to get hold of. As the molecular biologist Bob Holley found, this turned out to be the tRNA for alanine, since it ran differently on a chromatography column from all the others. Again the message to experimentalists is: Be sensible but don’t be impressed too much by negative arguments. If at all possible, try it and see what turns up. Theorists almost always dislike this sort of approach.
The path to success in theoretical biology is thus fraught with hazards. It is all too easy to make some plausible simplifying assumptions, do some elaborate mathematics that appear to give a rough fit with at least some experimental data, and think one has achieved something. The chance of such an approach doing anything useful, apart from soothing the theorist’s ego, is rather small, and especially so in biology. Moreover I have found, to my surprise, that most theorists do not appreciate the difference between a model and a demonstration, often mistaking the latter for the former.
In my terminology, a “demonstration” is a “don’t worry” theory (see the one described on page 97). That is, it does not pretend to approximate to the right answer, but it shows that at least a theory of that general type can be constructed. In a sense it is only an existence proof. Curiously enough, there exists in the literature an example of such a demonstration in relation to genes and DNA.
Lionel Penrose, who died in 1972, was a distinguished geneticist who in his later years held the prestigious Galton chair at University College, London. He was interested in the possible structure of the gene (which not all geneticists were at that time). He also loved doing “fretwork” (as it is called in England), making objects out of plywood with a fine saw. He constructed a number of such models to demonstrate how genes might replicate. The wooden parts had ingenious shapes, with hooks and other devices, so that when shaken they would come apart and join together in an amusing way. He published a scientific paper describing them and also a more popular article in
Scientific American.
An account by his son, Roger Penrose, the distinguished theoretical physicist and mathematician, appears in his father’s obituary written for The Royal Society.
I was taken to meet Lionel Penrose and his models by the zoologist Murdoch Mitchison. I tried to show a polite interest but had some difficulty in taking it all seriously. What to me was bizarre was that this was in the middle 1950s,
after
the publication of the DNA double helix. I tried to bring our model to Penrose’s attention but he was far more interested in his own “models.” He thought that perhaps they might be relevant for a pre-DNA period in the origin of life.
His wooden pieces, as far as I could see, had no obvious relation to known (or unknown) chemical compounds. I cannot believe that he thought genes were made of pieces of wood, yet he didn’t seem at all interested in organic chemicals as such. Why, then, was his approach of so little use? The reason is that his model did not approximate the real thing closely enough. Of course, any model is necessarily a simplification of some sort. Our DNA model was made of metal, but it embodied very closely the known distances between chemical atoms and, in the hydrogen bonds, took into account the different strengths of the various chemical bonds. The model did not itself obey the laws of quantum mechanics, but it embodied them to some extent. It did not vibrate, due to thermal motion, but we could make allowance for such vibrations. The crucial difference between our model and Penrose’s was that ours led to detailed predictions on matters that had not been explicitly put into the model. There is perhaps no precise dividing line between a demonstration and a model, but in this case the difference is very clear. The double helix, since it embodied detailed chemical features, was a true model, whereas Penrose’s was no better than a demonstration, a “don’t worry” theory.
It was all the more odd that his “model” came well after ours. What was its fascination for him? I think, at bottom, he
liked
to do fretwork, to play with little pieces of wood, and he was delighted that his favorite hobby could be used to illuminate one of the key problems in his professional life—the nature of the gene. I suspect that, on the other hand, he disliked chemistry and didn’t want to be bothered with it.
I cannot help thinking that so many of the “models” of the brain that are inflicted on us are mainly produced because their authors love playing with computers and writing computer programs and are simply carried away when a program produces a pretty result. They hardly seem to care whether the brain actually uses the devices incorporated in their “model.”