Eight Little Piggies (33 page)
Read Eight Little Piggies Online
Authors: Stephen Jay Gould


A plate of
Microdictyon
showing the characteristic meshwork. Acta Palaeontologica Sinica,
Vol. 28, No. 1, p. 5
.
In 1989, three Chinese colleagues from the Nanjing Institute of Geology and Paleontology—Chen Jun-yuan, Hou Xianguang, and Lu Haozhi—published a remarkable article in volume 28 of the
Acta Paleontologia Sinica
. (I remain profoundly grateful for the international character and cooperative traditions of paleontological work. Our science is global, and we would be stymied if we lost access to information from selected parts of the world. I thank both Drs. Chen and Hou for sending me reprints of their work along with letters providing further valuable data about their discoveries.)
Drs. Chen, Hou, and Lu have been working with the remarkable Chengjiang fauna of south-central China, an equivalent in age and soft-bodied preservation of the famous Burgess Shale in western Canada. Among other stunning creatures of unknown affinity, they discovered several specimens of a worm-like animal, some 8 cm in length.
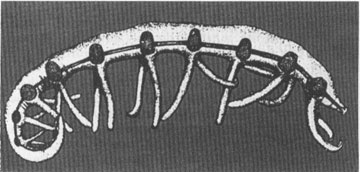
Restoration of
Microdictyon sinicum
showing the paired side plates. Acta Palaeontologica Sinica,
Vol. 28. No. 1, p. 5
.
This creature (see figure) bore ten thin pairs of leglike appendages, generally decreasing in strength from front to back. Traces of a simple, tubular gut can be seen on most specimens. But, most remarkable of all, this animal carried pairs of rounded phosphatic caps, inserted in pairs on the body sides, just above the joining points of the legs with the trunk. Each pair of legs, in other words, sports a corresponding pair of caps on the trunk above. These caps,
mirabile dictu
, are the elements previously named
Microdictyon
, but known only from the uninterpretable hard-part dabs. Their discovery on the Chinese animal not only adds a fascinating and mysterious creature to the roster of earliest animals, but also confirms our long-held suspicions about the SSF.
Microdictyon
, at least, is just an element covering a much larger body. Moreover, the hard parts enclose only a small portion of the body and do not articulate with each other (the space between pairs of caps is about double the diameter of the caps themselves). How could we possibly have inferred the character of the animal from the caps alone?
Halkieria
forms an even better known and more frustrating element of the SSF. Collected from lower Cambrian rocks throughout the world, halkieriids are preserved as sclerites (flattened blades and spines of calcification, just a millimeter or two in length). The sclerites assume several characteristic shapes, named siculate (narrow, crescentric, and asymmetrical), cultrate (elongate and more symmetrical), and palmate (wider and flattened like the palm of a hand). Although some paleontologists have tried to reconstruct halkieriids as tiny creatures, each living within or around a single sclerite, most agree that the halkieriid animal must have been substantially bigger and covered with large numbers of sclerites.
In July 1989, in a most inhospitable spot on Peary Land in northern Greenland, some twenty-one specimens of a halkieriid animal were finally unearthed from another deposit capable of preserving soft parts. (The spectacular results from the Burgess Shale have inspired paleontologists to devote attention to the discovery and exploitation of these rare and precious soft-bodied fossil faunas. Science, at its best, not only answers questions, but provokes new problems and guides fruitful research by posing issues previously unconsidered.)
In July 1990, S. Conway Morris and J. S. Peel published the first report on the halkieriid animal. Again, paleontological suspicions are confirmed, but with an amazing twist and surprise. The halkieriid is, as anticipated, a large animal (up to 7 cm), bearing many sclerites—up to 2,000 or more. The body is elongate, flattened, and wormlike, with the sclerites arranged in zones corresponding to forms previously named (see figure). Siculate sclerites surround the base of the animal (the underlying bottom surface probably carried no hard parts, as the animal crawled on a naked sole). A groove separates the zone of siculate sclerites from a lateral region of cultrates. The top surface of the creature bears more flattened, palmate sclerites.
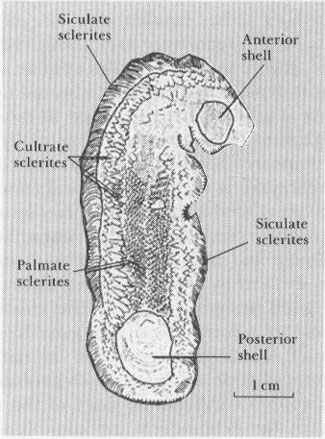
Figure of a halkieriid showing the fields of sclerites and the two end plates.
Reprinted by permission from
Nature,
June, p. 808; Copyright © 1990 Macmillian Magazines Limited
.
So far, so good—and quite in line with predictions. But nature always throws us a surprise or two. Each end carries a prominent and entirely unanticipated shell. These are found in the same position on every specimen, and therefore represent no fluke of juxtaposition or odd preservation. The anterior shell is roughly rectangular, the larger posterior shell (up to 1 cm in length) more oval and flattened. With apparent growth lines and an apex near the margin, this posterior shell, if found separately, would surely have been called a brachiopod or mollusk valve. (I suspect that several named mollusks and brachiopods of the Tommotian will turn out to be halkieriid end plates). Conway Morris and Peel wisely offer no interpretation of these elements, though others have suggested that the terminal shells might have plugged the ends of a U-shaped tube, if halkieriids burrowed as do many modern worm-shaped organisms.
We are, of course, enormously gratified to know, for the first time, some prominent animals of the SSF fauna, the earth’s initial complement of modern creatures with hard parts. But should we be surprised? (I realize that the phrase “SSF fauna” is as redundant as pizza pie and AC current, but abbreviations achieve a life of their own and may then be modified, even by one of their own elements).
One misguided reply might proffer little surprise (and relative indifference to my efforts in this essay). After all, we knew that the scattered SSF elements had to represent some kind of animal or other, and now that we have found two of the creatures, they turn out to be something rather familiar after all. Just a couple of worms—and as Mr. Reagan once said about redwoods, when you’ve seen one, you’ve seen ’em all. But such an attitude would be more than just deplorably Philistine; it would be dead wrong as well.
Wormlike
is a functional term used to describe flexible, soft-bodied organisms that are basically bilaterally symmetrical, with sensory organs in a head at front, and excretory organs at the rear end. Wormlike is not a genealogical concept uniting a group of organisms related in any evolutionary sense of common parentage.
Microdictyon
and
Halkieria
are wormlike only in this functional meaning, and no anatomical plan is more common and more often evolved by radically different creatures. Wormlike bodies are good designs for any mobile creature that must move with efficiency towards food and away from enemies—and no mode of life is more common in nature. Modern wormlike creatures include animals of such genealogical disparity as truly segmented earthworms, slugs of the snail lineage, sea cucumbers of the echinoderm phylum,
Amphioxus
of our own parentage (or at least cousinship), and a host of phyla that we all once learned in high school—Platyhelminthes (including laboratory planaria and tapeworms in vertebrate intestines), Nematoda, Kinorhynchia, Pogonophora, Chaetognatha, and so forth.
The proper evolutionary perspective is genealogical. Bats may be functionally similar to birds, but they are mammals by descent. Ichthyosaurs may look and work like fishes, but they are reptiles by ancestry. In this more fundamental context of genealogy, both
Microdictyon
and
Halkieria
are puzzling. The
Microdictyon
animal looks like an onychophoran, a small modern group considered by some as transitional between the Annelida (segmented worms) and the Arthropoda (insects, spiders and crustaceans—see next essay).
Halkieria
has been compared with the later
Wiwaxia
from the Burgess Shale, but
Wiwaxia
itself is an enigma, and the two shells at the end of
Halkieria
are just plain odd. Perhaps better evidence will establish some homologies with known groups, but for now,
Halkieria
must be viewed as a unique creature of unknown affinity with any other animal.
Thus, we may dismiss the “seen one worm, seen ’em all” argument as simply wrong, but a more sophisticated version of “should we be surprised” does have potential merit. Consider any genealogical system that ends up with a few well-differentiated survivors, all rather distant one from the other. Modern life surely displays this cardinal feature. Our modern phyla represent designs of great distinctness, and our diverse world contains nothing in between sponges, corals, insects, snails, sea urchins, and fishes (to choose standard representatives of the most prominent phyla). A distant past must have included many linking forms, now extinct. These links would not resemble fanciful hybrids between living organisms (a cat-dog or a cow-horse), because modern lineages have been separate for so long. They would, instead, be odd animals with veiled hints of several lineages to come and many unique features of their own (as we actually find in mammals like
Hyracotherium
, the 50-million-year-old ancestor of both horses and rhinoceroses).
Consider a figure and a nonbiological analogy (with thanks to R. T. Simmonds of Nordland, Washington, who wrote to me about this example in another context). The modern Romance languages—French, Spanish, Portuguese, Italian, and Romanian—all derive from Latin and represent clearly separate entities, despite evident similarities. But if we could—as we cannot—trace all the lineages leading from Latin, we would find a forest of village dialects linking all these end-points together. Many would be odd and unique, others smoothly transitional. We would learn that our modern descendants are just a small sample of the total richness, most now lost. We do get some hints of the full tree in survival of a few “minor phyla” (Catalan and Romansh, for example), and in historical records of a few extinct lineages (Provençal and Burgundian). But if we could go back to the beginnings of the spread, Dr. Simmonds conjectures, we would probably encounter a veritable Cambrian Explosion of lost variants.
In this sense, a phenomenon like the Cambrian Explosion must generate a majority of lineages that will seem peculiar in comparison with modern survivors—for these form the web of intermediary links that must die out if we are to emerge (as we have) with a limited set of widely separated designs (see figure). But I would raise two strong arguments against any boredom about
Microdictyon
and
Halkieria
on these grounds.
First of all, the forest of extinct lineages includes two categories of differing degrees of strangeness with respect to modern survivors. Unless modern survivors include forms at all the ancient peripheries—and this seems most unlikely, since peripheries are tenuous places—then many extinct lines will lie outside the range of all modern designs, and will feature more than an amalgam of primitive, but intermediary, characters. The point may sound abstract, but can be easily grasped in the diagram. Only lineages 1, 2, and 3 have emerged from the forest of this “Cambrian Explosion” to yield modern descendants. Now consider the lettered representatives from an early time of maximal diversity. Some of these lineages (e-l of the diagram) do lie within the bounds of modern groups; in our retrospective view, we will regard them as unique, but not fetchingly odd. But other lineages (a-d and m-p of the diagram) lie outside the limits of modern groups, often well beyond (p, for example, lies further from lineage 3 than 3 does from any modern survivor). These creatures will be read in our parochial light (recognizing only 1, 2, and 3 for “standard” animals) as bewitchingly peculiar—and all but the most benighted dolt will take a keen interest. (The Burgess Shale excites our imagination largely because several of its “weird wonders” probably lie in this exterior domain, well outside the boundaries of modern groups).