Eight Little Piggies (6 page)
Read Eight Little Piggies Online
Authors: Stephen Jay Gould

But why did
Lottia
gain the dubious honor of first marine invertebrate to disappear during historic time? Carlton and coauthors have also provided a coherent and satisfying explanation that neatly combines a specific historical event with the general biology of
Lottia
. Between 1930 and 1933,
Zostera
virtually disappeared from both the eastern and western Atlantic Ocean. (This species of sea grass has suffered numerous declines throughout its recorded history, but none nearly so severe as this accidental correlate with economic depression on adjacent lands.) The cause of this near wipe-out has been debated for years, with disease and environmental fluctuation as leading contenders. A series of articles published during the 1980s has decisively implicated a marine protist, the slime mold
Labyrinthula
(a unicelled creature that aggregates to form temporary colonies), as chief culprit. Infestation by the pathogenic species of
Labyrinthula
leads to formation of small black patches on
Zostera
leaves. The patches spread, eventually causing death and detachment of the entire blade.
This massive die-off of
Zostera
led to marked changes in associated ecosystems, including great reduction in migratory waterfowl populations and loss of commercial scallop fisheries. But neither
Zostera
, nor any of these ecological dependents, became entirely extinct.
Zostera
itself tolerates a much wider range of salinities than does the pathogen
Labyrinthula
. All populations of normal marine salinity were completely destroyed, but
Zostera
can also live in brackish water, while
Labyrinthula
cannot. Thus, relict
Zostera
populations hunkered down in low salinity refugia, and the species survived. Other species associated with
Zostera
also pulled through, either because they could also tolerate the low salinities of
Zostera
refuges, or because they could live on other resources, though often with much smaller populations, while
Zostera
was absent. When
Zostera
returned after the
Labyrinthula
epidemic subsided, these other species reflowered as well.
But consider the cruel fate of poor
Lottia alveus
. This limpet lacked the flexibility of all other species associated with
Zostera. Lottia
could not hunker down with
Zostera
in the low-salinity refugia because this limpet could only live in normal marine waters. And
Lottia
could not switch to another host species because it ate only the epithelial cells of
Zostera
blades. For
Lottia
, the total disappearance of
Zostera
in waters of normal salinity spelled complete destruction.
Zostera
returned, but no
Lottia
remained to greet the renewed bounty.
Does the story of
Zostera
and
Lottia
bear a message for our chief parochial concern with the subject of extinction—anthropogenic assault on the biosphere and consequent loss of biodiversity? In a literal sense, the answer must be “rather little.”
Lottia
was no Galapagos tortoise or passenger pigeon—creatures hounded to death by human hunters.
Lottia
didn’t even fall victim to some unintended consequence of human disruption in natural habitats. In fact,
Homo sapiens
probably played no role whatsoever in the death of
Lottia
(I doubt that one person in a million ever laid eyes on the creature).
Lottia
’s extinction was an ordinary natural event, the kind that, summed through geological ages, produces the basic pattern of life’s history. Epidemics are as intrinsic as water and sunshine in the history of life. They don’t usually wipe out a species entirely (as the
Labyrinthula
epidemic spared
Zostera
). Yet species clearly have differential susceptibility to extinction, and some factors of weakness enhance the possibility of death in epidemics. Natural selection can only work for immediate reproductive advantages and cannot overtly protect a species against unexpected vicissitudes of time. Many strengths of the moment engender a potential for later extinction as an unintended and detrimental side consequence. So long as
Zostera
bloomed and seas stood at normal salinity, intense specialization may have aided individual
Lottia
. But such narrowly committed forms are usually the first to go when unusual circumstances wipe out a highly specific habitat, even temporarily. Marine species are “relatively immune” from extinction because few have such narrow commitments, but
Lottia
, as an exception for its intense specialization, paid the ultimate price.
Lottia
does bear a symbolic message for the anthropogenic theme, however. As the first species to die (during historic times) in the one habitat that, from Lamarck to 1991, seemed free of such danger,
Lottia
must stand as a warning and an emblem—as the Crispus Attucks of a potential wave in the most protected arena, if our environmental assaults worsen. Didn’t British power laugh at the ragtag rebellion when Attucks and four others died in the Boston Massacre of 1770? Most crises start with something small, something virtually beneath notice. But whispers soon grow to whirlwinds. Limpets, with their low profiles and large apertures (often serving as suction cups for attachment), are metaphors for tenaciousness, for hunkering down in times of trouble. How appropriate, then, as a warning against complacency, that a real version of this symbol should be the first species to die in a realm of supposed invulnerability.
RICHARD OWEN
, England’s greatest vertebrate anatomist during Darwin’s generation (see Essay 5), developed the concept of an archetype to explicate the evident similarities that join us with frogs, flamingoes, and fishes. (An archetype is an abstract model constructed to generate an entire range of anatomical design by simple transformation of the all-inclusive prototype.) Owen was so pleased with his conception that he even drew a picture of his archetype, engraved it upon a seal for his personal emblem, and, in 1852, wrote a letter to his sister Maria, trying to explain this arcane concept in layperson’s terms:
It represents the archetype, or primal pattern—what Plato would have called the “divine idea” on which the osseous frame of all vertebrate animals—i.e. all animals that have bones—has been constructed. The motto is “the one in the manifold,” expressive of the unity of plan which may be traced through all the modifications of the pattern, by which it is adapted to the very habits and modes of life of fishes, reptiles, birds, beasts, and human kind.
Darwin took a much more worldly view of the concept, substituting a flesh and blood ancestor for a Platonic abstraction from the realm of ideas. Vertebrates had a unified architecture, Darwin argued, because they all evolved from a common ancestor. The similar shapes and positions of bones record the historical happenstance of ancestral form, retained by inheritance in all later species of the lineage, not the abstract perfection of an ideal shape in God’s realm of ideas. Darwin burst Owen’s bubble with a marginal note in his personal copy of Owen’s major work,
On the Nature of Limbs
. Darwin wrote: “I look at Owen’s archetype as more than idea, as a real representation as far as the most consummate skill and loftiest generalization can represent the parent form of the Vertebrata.”
However we construe the concept of an organizing principle of design for major branches of the evolutionary tree—and Darwin’s version gets the modern nod over Owen’s—the idea remains central to biology. Consider the subset of terrestrial vertebrates, a group technically called Tetrapoda, or “four-legged” (and including amphibians, reptiles, birds, and mammals in conventional classifications). Some fly, some swim, and others slither. In external appearance and functional role, a whale and a hummingbird seem sufficiently disparate to warrant ultimate separation. Yet we unite them by skeletal characters common to all tetrapods, features that set our modern concept of an archetype. Above all, the archetypal tetrapod has four limbs, each with five digits—the so-called pentadactyl (or “five-fingered”) limb.
The concept of an archetype does not require that each actual vertebrate display all canonical features, but only that uniqueness be recognized as extreme transformations of the primal form. Thus, a whale may retain but the tiniest vestige of a femur, only a few millimeters in length and entirely invisible on its streamlined exterior, to remind us of the ancestral hind limbs. And although a hummingbird grows only three toes on its feet, a study of embryological development marks them as digits two, three, and four of the full ancestral complement. The canonical elements are starting points and generating patterns, not universal presences.
In the tetrapod archetype, no feature has been more generally accepted than the pentadactyl limb, putative source of so many deep and transient human activities, from piano playing to touch typing, duck shooting, celebratory “high fives,” and decimal counting (twice through the sequence of “this little piggy…”). Yet this essay will challenge the usual view of such a canonical number, while not denying its sway in our lives.
The great Swedish paleontologist, Erik Jarvik, closed his two-volume magnum opus on vertebrate structure and evolution with a telling point about pentadactyl limbs and human possibilities. He noted how many “advanced” mammals modify the original pattern by loss and specialization of digits—horses retain but one as a hoof; whales practically lose the whole hind limb. Jarvik noted that an essential coupling of a multidigited hand, fit for using tools, with an enlarging brain, well suited to devising new and better uses for such technology, established the basis and possibility of human evolution. If the ancestor of our lineage had lost the original flexibility of the “primitive” pentadactyl limb and evolved some modern and specialized reduction, human intelligence would never have developed. In this important sense, we are here because our ancestors retained the full archetypal complement of five and had not substituted some new-fangled, but ultimately more limiting, configuration. Jarvik writes:
The most prominent feature of man is no doubt his large and elaborate brain. However, this big brain would certainly never have arisen—and what purpose would it have served—if our arm and hand had become specialized as strongly as has, for instance, the foreleg of a horse or the wing of a bird. It is the remarkable fact that it is the primitive condition, inherited from our osteolepiform ancestors [fishes immediately ancestral to tetrapods] and retained with relatively small changes in our arm and hand, that has paved the way for the emergence of man. We can say, with some justification, that it was when the basic pattern of our five-fingered hand for some unaccountable reason was laid down in the ancestors of the osteolepiforms that the prerequisite for the origin of man and the human culture arose.
I don’t dispute Jarvik’s general point: The retention of “primitive” flexibility is often a key to evolutionary novelty and radiation. But is the five-fingered limb a constant and universal tetrapod archetype, interpreted in Darwin’s evolutionary way as an ancestral pattern retained in all descendant lineages?
Erik Jarvik is maximally qualified to address this question (his rationale, of course, for raising it in the first place), for he has done by far the most extensive and important research on the earliest fossil tetrapods—the bearers and perpetrators of the five-fingered archetype in any evolutionary interpretation. (Fish fins are constructed on different principles, although the lobe-finned ancestors of tetrapods built a bony architecture easily transformable to the fore and hind limbs of terrestrial vertebrates. In any case, fish do not display the pentadactyl pattern, and this central feature of canonical design arose only with the evolution of the Tetrapoda.)
The oldest tetrapods were discovered in eastern Greenland by a Danish expedition in 1929. They date from the very last phase of the Devonian period, a geological interval (some 390 to 340 million years ago) often dubbed the “age of fishes” in books and museum exhibits that follow the silly chauvinism of naming time for whatever vertebrate happened to be most prominent. The Swedish paleontologist Gunnar Säve-Söderbergh collected more extensive material in 1931 and directed the project until his untimely death in 1948. Erik Jarvik then took over the project and, during the 1950s, published his extensive anatomical studies of two genera that share the spotlight of greatest age for tetrapods—
Ichthyostega
and
Acanthostega
. Although no specimens preserved enough of the fingers or toes for an unambiguous count, Jarvik (see figure) reconstructed the earliest tetrapods with the canonical number of five digits per limb.
The standard reconstruction of
Ichthyostega
from Jarvik’s 1980 book. Note the five digits on each limb.
From
Basic Structure and Evolution of Vertebrates,
vol. 1, p. 235
.
Our confidence in this evidence-free assumption of an initial five began to crumble in 1984, when the Soviet paleontologist O. A. Lebedev reported that the newly discovered early tetrapod
Tulerpeton
, also of latest Devonian age, bore six digits on its limbs. This find led anatomist and embryologist J. R. Hinchliffe to suggest in 1989, prophetically as we have just learned, that five digits represents a secondary stabilization, not an original state. Hinchliffe entitled his article “Reconstructing the archetype: Evolution of the pentadactyl limb,” and ended with these words: “Restriction to the pentadactyl form may have followed an evolutionary experimental phase.”
Hinchliffe’s suspicion has now been confirmed—in spades. In October 1990, M. I. Coates and J. A. Clack reported on new material of
Ichthyostega
and
Acanthostega
, collected by a joint Cambridge-Copenhagen expedition to East Greenland in 1987 (see bibliography). Some remarkable new specimens—a complete hindlimb of
Ichthyostega
and a forelimb of
Acanthostega
—permit direct counting of digits for the first time.
In an admirable convention of scientific writing that maximizes praise for past work done well and minimizes the disturbing impact of novelty, Coates and Clack write: “The proximal region [closest to the body] of the hindlimb of
Ichthyostega
corresponds closely with the published description, but the tarsus [foot] and digits differ.” In fact, the back legs of
Ichthyostega
bear, count ’em, seven toes!—with three smallish and closely bound digits corresponding to the hallux (“big toe” in human terms) of ordinary five-toed tetrapods (see figure).
Acanthostega
departs even more strongly from a model supposedly common to all; its forelimb bears eight digits in a broad arch of increasing and then decreasing size (see figure).
The conclusion seems inescapable, and an old “certainty” must be starkly reversed. Only three Devonian tetrapods are known. None has five toes. They bear, respectively, six, seven, and eight digits on their preserved limbs. Five is not a canonical, or archetypal, number of digits for tetrapods—at least not in the primary sense of “present from the beginning.” At best (for fans of pentadactyly) five is a later stabilization, not an initial condition.
Moreover, in the light of this new information, an old fact may cast further doubt on the primacy of five. The naive “ladder of life” view depicts vertebrate evolution as a linearly ascending series of amphibian—reptile—mammal—human (with birds as the only acknowledged branch). But ladders are culturally comforting fictions, and copious branching is the true stuff of evolution. Tetrapods had a common ancestor to be sure, but modern amphibians (frogs and salamanders) represent the termini of a large branch, not the inception of a series. Moreover, no fossil amphibian seems clearly ancestral to the lineage of fully terrestrial vertebrates (reptiles, birds, and mammals), called Amniota to honor the “amniote” egg (with hard covering and “internal pond”), the evolutionary invention that allowed, in our usual metaphors, “complete conquest of the land” or “true liberation from water.” (The point is tangential to this essay, but do pause for a moment and consider the biases inherent in such common “descriptions.” Why is the ability to lay eggs on land a “liberation” is water tantamount to slavery? Why is exclusive dwelling a “conquest”? Who is fighting for what? Such language only makes sense if life is struggling upward towards a human pinnacle—the silliest and most self-centered view of evolution that I can imagine.)
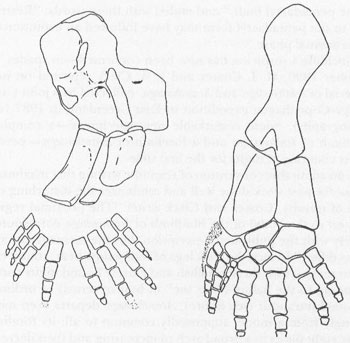
Left: forelimb of
Acanthostega
with 8 digits. Right: hindlimb of
Ichthyostega
with seven digits.
Reprinted by permission from
Nature
vol. 347, p. 67; Copyright © 1990 Macmillan Magazines Limited
.
The first fossil reptiles are just about as old as the first amphibians clearly in the group that eventually yielded our modern frogs and salamanders. Thus, rather than a ladder from amphibian to reptile, both the fossil record and the study of modern vertebrate anatomy suggest an early branching of the tetrapod trunk into two primary limbs—the Amphibia and the Amniota (reptile, bird, and mammal).
And now, the point about pentadactyly and its limits: The Amniota do, indeed, show the canonical pattern of five toes upon each limb (or some modification from this initial state). But Amphibia, both living and fossil, have five toes on the hindlimbs and
only four
on the front limbs. Anatomists have known this for years of course, but have always assumed that this reduction to four proceeded from an initial and canonical five. This conclusion must now be challenged. If all the earliest tetrapods had more than five digits, and if amphibians have been separate from amniotes since the beginning of terrestrial life, why assume that the four toes of the amphibian forelimb descended from a primary five? All modern stabilizations probably proceeded from more than five. Perhaps the amphibian forelimb went from this higher number directly to four, without any pentadactyl stage between. If so, then pentadactyly crumbles on two grounds: (1) It does not represent the original state of tetrapods (as six-, seven-, and eight-toed earliest forms show); and (2) it may not mark the canonical state in one of the two great living lineages of tetrapods.