Eight Little Piggies (9 page)
Read Eight Little Piggies Online
Authors: Stephen Jay Gould

The appearance in the tail of the
Ichthyosaurus
…is too uniform and common to be due entirely to an accidental and extrinsic cause. I am therefore disposed to attribute it to an influence connected with some structure of the recent animal; and most probably to the presence of a terminal…caudal fin, which, either by its weight, or by the force of the waves beating upon its extended surface, or by the action of predatory animals of strength sufficient to tug at without tearing it off, might…give rise to a dislocation of the caudal vertebrae immediately proximal to its attachment.
The puzzle finally achieved its solution in the 1890s when the perennial, but rarely granted, prayer of all paleontologists was answered by the powers that be. Ichthyosaurs with preserved soft parts were discovered in the Holzmaden deposits near Stuttgart. These sediments are so rich in organic oils and bitumen that they actually burn. (One fire raged beyond control from 1668 to 1674 and another from 1937 to 1939). Details of internal organs are not retained in these bitumen beds, but body outlines remain intact as black films upon the light gray rock. (Most of the fine specimens displayed at museums throughout the world come from the Holzmaden beds, and many readers are no doubt familiar with ichthyosaur body outlines preserved as blackened films on the rock under and behind the bones.)
The Holzmaden ichthyosaurs finally proved the extent of external convergence upon the stereotypical form of a free-swimming fish. The dorsal fin, with no bony support at all, was revealed for the first time. And the caudal fin, correctly inferred by Owen from the tailbend, now stood out for all to see. The fin was vertical, as Owen had surmised, and composed of two nearly equal and symmetrical lobes. The vertebral bend did mark, again as Owen had conjectured, the anterior border of the fin—but as an item of normal anatomy, not a postmortem artifact or dislocation. The vertebral column bent naturally down to follow the lower border of the lower lobe of the tail right to the animal’s rear end. No other vertebrate displays this orientation. In fishes, the vertebral column either stops at the inception of the tail or extends, as in sharks, into the upper border of the upper lobe. No wonder that the ichthyosaur tailbend had provoked such confusion for more than fifty years.

A tossil ichthyosaur with characteristic and excellent preservation from the Holzmaden deposits. Note the outlines of back and tail fins, and also the bending of the vertebral column into the lower lobe of the tail.
Courtesy of
Natural History.
Nearly a century has passed since the Holzmaden discoveries revealed the true nature of the ichthyosaur tailbend by exposing its enclosure within the caudal fin. Yet the tailbend continues to provoke commentary and controversy for two main reasons as outlined by Chris McGowan of the Royal Ontario Museum, Toronto, and the world’s leading expert on ichthyosaurs (my thanks to Dr. McGowan not only for his many illuminating articles, but especially for enduring a long phone call of inquiry during my research for this essay). First, and positively, the location, angle of downturn, and length of the vertebral column after the bend specify both the size and form of the caudal fin (only the Holzmaden ichthyosaurs preserve the fin itself as a carbonized film; all other specimens are bones alone, and the tailfin must be inferred from the vertebral column).
Second, and representing yet another dubious triumph of expectation over observation (perhaps the most common of human foibles), many classic specimens have been reconstructed on the assumption that tailbends must be present. I raise no issue of fraud or delusion. In many specimens, the vertebrae (particularly the small items at the rear end) lie scattered over the rock surface. The wonderfully expert and professional Holzmaden preparators adopted the custom of removing these bones entirely from the matrix and then resetting them in the inferred position of the living animal—that is, with a tailbend. We have no doubt that several ichthyosaur species developed a pronounced tailbend, since perfect specimens with preserved body outlines clearly show the tail vertebrae extending into the lower lobe of the caudal fin. But perhaps other species (particularly the earliest forms) lacked a tailbend, and perhaps preparators have tended to exaggerate the amount of inclination in reconstructing their specimens.
If the actual tailbends of most specimens on display are thus infected with doubt, how can we be confident about the existence and form of the caudal fin in most species? And, since this information is crucial to our understanding of swimming and maneuvering in ichthyosaurs, how can we hope to reconstruct the ecology of these fascinating animals? Obviously, we need a criterion of confirmation separate from the bend itself. Fortunately, McGowan has been able to establish such a criterion and to devise an ingenious way of putting it into practice.
How can an angular bend be produced in a basically linear structure (like the vertebral column), built from a sequence of disks that must follow, one behind the other, without large spaces between? As the accompanying sketch shows, tailbends imply a change in the shape of the crucial vertebral disks at the bend itself—from their usual form (with upper and lower borders of equal width) to a wedge with a wider border on top and a narrower edge below. A succession of wedge-shaped disks will inevitably cause the tail to bend, and the greater the difference in width between upper and lower borders, the more pronounced the bend. In fact, by a simple construction akin to the problems we all worked in high-school plane geometry, the angle of the bend can be inferred from the number of wedge-shaped disks and their intensity of wedging.
But how can this wedging be assessed? The vertebrae of most skeletons are at least partially embedded in rock, and both ends are rarely exposed to reveal the extent of wedging (while museums rarely look kindly upon requests for sufficient mayhem upon their specimens to dig the vertebrae out of the enclosing matrix). McGowan solved this problem with a boost from modern medical technology—computed tomography as provided by a CT-scanner. These marvelous, donut-shaped x-ray devices can take a photographic slice right through a human body in any orientation (so long as the body fits into the donut-hole of the machine). Well, an ichthyosaur in its matrix is often about the same size as a human body. Why not take a CT-scan of vertebrae at the tailbend, thus producing a photographic image of the vertebral disks while still embedded in their matrix? (McGowan didn’t initiate the application of CT-scanning to paleontological material. Several successful attempts have been made in the past few years, including the resolution of cranial capacities and form of unerupted teeth in some important skulls of the human fossil record.)
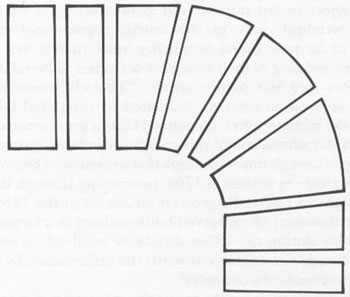
Shape of vertebral disks in ichthyosaurs with tail bends. Note the necessary wedge shape at the bend itself.
Ben Gamit
.
McGowan used a CT-scanner to affirm that
Leptopterygius tenuirostris
, an early ichthyosaur with an uncertain tailbend currently subject to hot dispute, did grow a series of six wedge-shaped vertebral disks in the crucial region—not strongly wedged to be sure (none producing more than a five degree bend), but yielding in their ensemble a modest tailbend of some 25 degrees (see McGowan’s article, “The ichthyosaurian tailbend: A verification problem facilitated by computed tomography,” in the bibliography). Somehow, I feel a great sense of satisfaction in the affirmation of this continuity in human striving for knowledge through time—to think that a discussion beginning in two Latin treatises written in 1708, proceeding through the keen observations of England’s greatest anatomist in the 1830s, and on to the discovery of preserved body outlines in a famous German locality during the 1890s should be resolved, as we begin our last decade’s countdown towards the millennium, by the latest device of medical machinery!
Yet, however satisfying the particular resolution, this tale (and tail) would convey no message or meaning (to those outside the tiny coterie of ichthyosaurian aficionados) if the problem of the ichthyosaur tailbend did not illuminate something central in evolutionary theory. Ichthyosaurs are most celebrated for their convergence upon the external form of superior swimmers among fishes. Since English traditions in natural history place primary emphasis on the concept of adaptation, these similarities of fish and marine reptile have won the lion’s share of written attention—for we know how the threefold combination of flippers, backfin, and tailfin work in efficient hydrodynamic coordination, and we are awed that two independent lineages evolved such uncanny resemblance for apparently similar function. This awe even predates evolutionary theory, for an earlier attribution to God’s benevolent care inspired as much admiration as our current respect for the power of natural selection. William Buckland, Owen’s close colleague, had a special affection for ichthyosaurs. He also wrote the greatest paean of the 1830s to adaptation as proof of God’s benevolence. In
Geology and Mineralogy Considered with Reference to Natural Theology
, written in 1836, Buckland invoked the precise convergence of ichthyosaur and fish as a proof of God’s goodness. Buckland acknowledged that an ordinary reptile would be in severe trouble at sea, but ichthyosaurs have been granted by divine fiat (read “endowed by natural selection” for a modern version of the same argument):
…a union of compensative contrivances, so similar in their relations, so identical in their objects, and so perfect in the adaptation of each subordinate part, to the harmony and perfection of the whole; that we cannot but recognize throughout them all, the workings of one and the same eternal principle of Wisdom and Intelligence, presiding from first to last over the total fabric of the Creation.
Yet, in our complex world of natural history, almost any profuse enthusiasm also elicits its mitigating opposite. (Such a cautionary splash of cold water may then emerge as a primary theme with more enlightening implications in itself.) Yes, ichthyosaur convergences are remarkable; only a soulless curmudgeon could fail to be impressed by the fishlike form of this descendant from ordinary terrestrial reptiles. Only the most militant denigrator of Darwin and the entire English tradition could fail to utter the word
adaptation
with both confidence and admiration.
But another perspective demands equal attention—and Owen, the much misunderstood proponent of a continental tradition that viewed adaptation as superficial, and sought regularities of form underneath a garb of immediate design, discussed ichthyosaurs primarily in the light of this alternative. What are the limits to adaptation imposed by the disparate anatomical designs underlying a convergence (fishes and reptiles in this case)? To what extent must the ichthyosaur remain in the thrall of its past, quite unable to mimic the form of a fish exactly because the historical legacy of a reptilian body plan precludes a large set of favorable options? To what degree, in short, must an ichthyosaur remain an easily identified reptile in marine drag?
For a primary statement of this alternate theme (limits imposed by inherited design), we must look to the largely forgotten work of the great Belgian paleontologist Louis Dollo (1857–1931). Dollo gave his name to an evolutionary principle known as
irreversibility
(often called Dollo’s Law). In one of the cruel ironies often imposed by history, many fine thinkers win their posthumous recognition only by eponymous linkage with a principle so widely misunderstood that true views turn into their opposite. Many evolutionists interpret Dollo’s Law as an antiquated statement about inherent, directional drives in evolution—a last gasp of a mystical vitalism that the Darwinian juggernaut finally defeated. In fact, Dollo was a convinced mechanist, and a Darwinian in basic orientation (with some interesting wrinkles of disagreement).
To Dollo, irreversibility epitomized the nature of history under simple conditions of mathematical probability (Dollo had obtained an extensive education in mathematics and attributed his formulation of irreversibility to this training). Evolutionary transformations are so complex—involving hundreds of independent changes—that any complete reversal to a former state becomes impossible for the same reason that you will never flip 1,000 heads in a row with an honest coin. No mysticism, no vitalism, only the ordinary operation of probability in a complex world. A simple change (increase in size, mutation in a single gene) may be reversed, but the standard transformations that form the bread and butter of paleontology (origin of flight in birds, evolution of humans from apelike ancestors) cannot run backwards to recover an ancestral state exactly.