Secondary Schizophrenia (126 page)
Read Secondary Schizophrenia Online
Authors: Perminder S. Sachdev

population.
316
Chapter 24 – Velocardiofacial syndrome
availability of dopamine in the prefrontal cortex
[130].
The prefrontal cortex is the seat of executive functioning and sustained attention and is contributory
to working memory. Increased dopamine levels associated with the Met allele are thought to confer a
cognitive advantage. In studies of healthy individuals, as well as patients with schizophrenia and schizotypy (unrelated to VCFS), individuals homozygous
for the Met allele have been shown to perform better on tests of prefrontal cognition, as compared to
those with Val homozygosity or heterozygosity for
Met/Val
[131, 132, 133].
Furthermore the Val allele
occurred often in individuals with schizophrenia spectrum disorders
[134, 135, 136].
Thus, there is considerable evidence that the Val/Met polymorphism is
associated with the neurocognitive deficits observed
in schizophrenia in the general population, especially
those related to executive function and working memory, although some studies have been contradictory
[137].
In individuals with VCFS, there is controversy about
the relationship between the COMT genotype, the cognitive deficits, and schizophrenia. Initial studies that
examined the association between the COMT polymorphism and neurocognition in children with VCFS
found that children hemizygous for the low activity
Met allele performed better on measures of executive function than Val hemizygous individuals
[138,
139],
a finding that was consistent with the reports in
the general population with and without schizophrenia as discussed previously. Other studies have found
that the Met allele is associated with poor neurocognitive functioning
[66],
minor psychiatric diagnoses,
such as ADHD
[140],
and may predict longitudinal
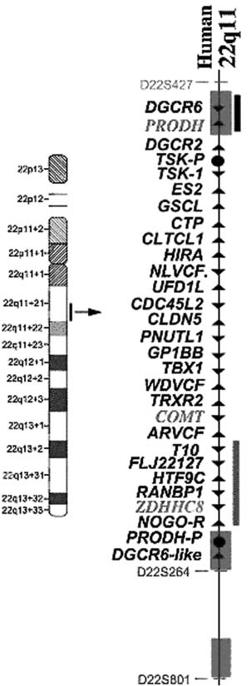
Figure 24.5
Ideogram of Chromosome 22, with a listing of all the
decline in verbal IQ as well as psychosis
[141].
In a
genes in the 1.5 Mb interval in the q11.2 region, thought to be
critical for schizophrenia. The genes denoted in red are thought to
study of adults with VCFS, there was no association
be most likely related to psychosis. (See color plate section.)
between the COMT genotype and schizophrenia
[10].
A new study of adults with VCFS reported that the
Met allele was associated with poor neurocognition,
COMT
but the Val/Met polymorphism was not associated with
The COMT (catechol-O-methyl transferase) protein is
schizophrenia
[142].
This apparent contradiction in
the major enzyme responsible for the degradation of
findings may in part be due to small sample sizes, dif-dopamine in the prefrontal cortex
[127],
unlike the
ferent age groups, as well as issues related to methodol-striatum, where degradation of dopamine is largely
ogy (selection of neurocognitive tests versus diagnos-due to the dopamine transporter gene
[128].
A com-tic psychiatric assessments) across the various studies.
mon SNP in the COMT gene, with the substitution
Also plausible is an unifying explanation that may help
of methionine (Met) for valine (Val) at codon 158,
explain the controversial reports of the effects of the
causes the Met allele to have one fourth the enzymatic
Val/Met polymorphism in VCFS: the concept that any
activity of the Val allele
[129]
, resulting in increased
deviation from the normal metabolism of dopamine
317
Organic Syndromes of Schizophrenia – Section 3
in the form of excessive or defective clearance is likely
work is needed before the role of PRODH in glu-to result in abnormal neurocognition, similar to that
taminergic transmission is clear. The PRODH gene
seen in schizophrenia
[143]
. However, given the rel-has also been implicated in apoptosis by supporting
atively recent evidence that the cognitive deficits in
the generation of reactive oxygen species
[155],
but
schizophrenia are indeed associated with a hypoactive
it is unclear if this property is related to schizophre-dopaminergic system in the prefrontal cortex as would
nia. Examination of this function of PRODH and
be expected with the Val allele, further exploration of
schizophrenia would be intriguing, because excessive
this issue in children with VCFS may yield important
apoptosis has been proposed as an additional mech-information
[144, 145, 146].
anism in the pathogenesis of the disorder
[25].
Sev-Other SNPs in the COMT gene have been ana-eral SNPs in the PRODH gene (rs2238731, rs2904551,
lyzed to determine a relationship to schizophrenia in
rs3970559, rs450046) have been reported to be associ-the general population and of interest are rs737865
ated with schizophrenia in the general population, of
and rs165599
[147, 148].
The three marker hap-which some are thought to be responsible for elevated
lotype consisting of genotypes at rs4680-rs737865-proline levels
[124, 126].
Thus, PRODH is regarded
rs165599 (rs4680
=
Val/Met polymorphism) is said
as an important candidate gene for schizophrenia,
to be associated with schizophrenia in the general
although not all studies have found such a link
[156,
population
[149].
As yet, the impact of such hap-
lotypes in individuals with VCFS has not been well
In individuals with VCFS, there are not enough
delineated.
studies on PRODH thus far; a small study involving
genotyping of two polymorphisms within the PRODH
gene did not find any specific associations between
PRODH
these and psychiatric diagnoses
[141].
Another study
The proline dehydrogenase (PRODH) gene product is
reported no association between the PRODH polymor-responsible for the degradation of proline, an amino
phism and schizophrenia in 12 patients with VCFS
acid that functions as an inhibitory neurotransmit-and schizophrenia
[158]
. A recent study reported that
ter and/or a metabolic precursor of glutamate
[150,
hyperprolinemia was found in 37% of individuals with
151].
Proline enhances glutamate release, resulting in
VCFS and that elevated proline levels were associated
increased synaptic transmission. Reduced glutamin-with lower IQ but not psychosis. Interestingly, these
ergic neurotransmission through the N-methyl-D-authors reported that elevated proline in combination
aspartate (NMDA) receptor, which is found in high
with the Met allele in the COMT gene was associated
concentrations in the prefrontal cortex, is believed to
with psychotic illnesses
[159].
Thus, the PRODH and
be related to schizophrenia, especially the deficits in
COMT genes in conjunction may modify psychosis
working memory
[152, 153].
A model implicating glu-risk, speculatively through the interaction between
tamate in the pathogenesis of schizophrenia proposes
dopaminergic and glutaminergic transmission, as has
that reduced glutamate during the prenatal and ado-been suggested in schizophrenia in the general popu-lescent periods accounts for the premorbid neurocog-lation
[128].
nitive manifestations and the onset of illness, because
reduced glutamate is believed to result in excessive
synaptic elimination. After the onset of the illness,
ZDHHC8
an increase in glutamate release occurs, secondary
This gene lies at the distal end of the critical deleted
to elevated dopamine levels, causing increased neu-segment and is involved in palmitoylation, a process
rotoxic cell loss, thus leading to disease progression
that consists of modification of proteins with the lipid
[25].
In a mouse model with PRODH deficiency, loco-palmitate. ZDHHC8 is highly expressed in the human
motor deficits were attributed to elevated proline lev-brain
[125]
and is thought to be important for synap-els enhancing glutaminergic transmission
[154].
These
tic transmission, because palmitoylation is believed to
authors did not find any deficits in spatial working
modify neurotransmitters as well as other neuronal
memory in PRODH deficient mice. It is possible that
proteins
[160].
A particular SNP rs175174 within the
similar to dopamine, deficiency and excess of gluta-gene results in the production of a truncated protein
mate may have different effects in different areas of
and has been demonstrated to have an association with
318
the brain at different stages of psychosis and further
schizophrenia
[125,
161]
. There are no studies of an
Chapter 24 – Velocardiofacial syndrome
association between this gene and schizophrenia in
Mouse models of VCFS: relevance to
humans with VCFS.
schizophrenia
Although the primary motive in creating a mouse
UFD1L
model of the 22q11.2 deletion was to characterize the
structural defects and the specific genes responsible for
The ubiquitin fusion degradation 1 gene is believed
these, there has been considerable interest in deter-to be important in embryogenesis. It is expressed in
mining the gene(s) that contribute to the psychiatric
the medial telencephalon, which forms the hippocam-manifestations as well. A major challenge in using
pus, in fetal life starting at 10 weeks of gestation
the mouse model for this purpose is that it is diffi-and continuing into postnatal life. A functional SNP
cult to identify core human psychiatric traits that can
rs5992403 that results in upregulation of the gene has
be effectively studied in mice. Indeed, this will con-been studied and has been found to be associated with
tinue to pose limitations to the amount of information
schizophrenia in the general population
[162].
It is
pertinent to schizophrenia that can be gleaned from
postulated that both upregulation and downregula-the mouse model. This being said, traits that can be
tion of the gene (the latter would be true of VCFS) is
characterized are: anxiety, decreased sociability, and
believed to result in neuronal dysfunction and apop-impaired working memory and sensorimotor gating
tosis
[163]
, but, thus far, there have been no reports of
(measured by prepulse inhibition). The most studied
SNPs in this gene being linked to the cognitive or psy-of these is the prepulse inhibition (PPI) of the acous-chiatric abnormalities in individuals with VCFS.
tic startle reflex, considered a phenotypic marker for
schizophrenia, although not a specific one, because it
is also seen in obsessive-compulsive disorder, Tourette
TBX1
syndrome, and Asperger syndrome
[167].
One of several transcription factors involved in devel-Various knockout mouse models have been uti-opmental processes, the TBX1 gene, has been shown
lized to study the genes responsible for the neuropsy-to be responsible for several of the phenotypic fea-chiatric manifestations in VCFS. A region of the mouse
tures of VCFS, including the cardiac anomalies, facial
chromosome 16 is homologous to the 22q11.2 interval
features, thymic hypoplasia, and the parathyroid dys-in humans: the models created thus far include mice
function
[164].
Indirect evidence for a possible role of
with heterozygous deletions of genes in the interval-TBX1 in the neurobehavioral manifestations of VCFS
(Dfl/
+
)
[168]
and Lgdel/
+
[169],
as well as mice mod-comes in the form of abnormal sensorimotor gating in
els with specific single gene mutations, as described
mice models who are haploinsufficient for TBX1
[165].