The Flamingo’s Smile (8 page)
Read The Flamingo’s Smile Online
Authors: Stephen Jay Gould

In the first pages of his monograph, Serres tries to reduce the embryology of all animals to three basic laws of “organology.” First, by the law of
eccentric development
, known otherwise as the law of circumference to center, organs form initially at the edge of the developing embryo and then migrate toward the center. Second, by the law of
symmetry
, organs that become single and central in adults begin as double symmetrical rudiments on opposite edges of the developing embryo. Third, by the law of
affinity
, these symmetrical rudiments are drawn towards each other until they fuse in the center to form a single adult organ. (Let me be charitable and simply state that these laws are unwarranted extensions of patterns that operate occasionally in development. Serres was writing before the establishment of cell theory and just a few years after Karl Ernst von Baer’s discovery of the human ovum. His formal approach to morphology, so foreign to a world that can assess cellular and even molecular causes, fit the knowledge and mores of his own era.)
Two hundred pages later, when Serres finally discusses his dissection of Ritta-Christina, we understand why he devoted so much preceding space to the three primary laws of organology—for they provide his solution to the great dilemma of individuality. Ritta and Christina are two people, albeit imperfect, and the laws of form proclaim
their
status.
No one quarreled with the double verdict on Ritta and Christina from the waist up; the dilemma had always centered upon their well-formed, but clearly single, lower half—one anus, one vaginal opening, two legs. If she were two people all the way from stem to stern, how could her lower half form so well in the shape of one? How could the incomplete parts of two separate creatures fuse and blend into a form indistinguishable from the lower half of such unambiguous singletons as you or me?
Serres used his laws of organology to render Ritta and Christina’s lower half as the conjoined product of two people. After all, the harmonious, well-formed single organs of ordinary individuals arise (by the law of symmetry) as separate and double parts at the embryo’s edge, and then move inward (by the law of circumference to center), eventually meeting and fusing (by the law of affinity) into one integral organ. If our single heart, stomach, and liver begin as two symmetrical rudiments (actually, they don’t, but Serres thought they did), then why should we view the presence of a single, well-formed organ in Ritta and Christina’s lower half as any argument against its construction from the mingled and melded parts of two embryonic individuals? If the twins have but one uterus, then the right half came from Ritta, the left from Christina. The two rudiments formed at the embryonic edges, in regions unambiguously assigned to Ritta or to Christina (law of symmetry). They moved toward the midline (circumference to center) and joined there (law of affinity) to form a single organ.
Serres announced proudly that his laws of form had resolved the great dilemma in favor of duality: “How could we possibly have conceived that each child furnished half of an organ common to both, if the law of eccentric development had not taught us that single organs are, in their normal state, originally double.”
Serres did not shrink from the decidedly peculiar logical implications of his solution. He noted that the large uterus had proper connections with the ovaries and vaginal canal and saw no reason why Ritta and Christina might not have borne children had they lived to maturity. (Serres also found a second, rudimentary uterus that would not have worked.) He concluded that the large uterus had formed half from Ritta and half from Christina, and admitted that any offspring developed within would have two natural mothers:
This disposition of Ritta and Christina’s genital organs evidently shows…that while nature had taken measures to assure the lives of these children, she had not forgotten the possibility of their reproduction. Now, for this reproduction, nature had combined everything, so that all the pleasures and pains would be shared…. Supposing that conception occurred in the large uterus, a single child would have had two distinct mothers, a singular result of this associated life.
Serres then discussed a pair of conjoined males with four legs and a single head, and opted for consistency and duality: the single well-formed brain shared the combined thoughts of two.
There is a perfect unity produced by two distinct individualities. There are sense organs and cerebral hemispheres for a single individual, adapted to the service of two, since it is evident that there are two
me
’s in this single head [
deux
moi
dans cette tête unique
].
Thus Serres made a valiant and consistent attempt to resolve a question that seemed hopelessly ambiguous. We may appreciate the effort and enjoy an excursion into the different view of biology that Serres maintained. But we must reject his conclusion.
Fertilized human eggs usually develop into single individuals. Rarely, the dividing cells separate into discrete groups, and two embryos develop. These one-egg (or identical) twins are genetic carbon copies. In some ultimate, biological sense, they are the same iterated individual—and the psychological literature contains ample testimony to feelings of imperfect separation shared by many so-called identical twins. Yet, at least for definition’s sake, we experience no difficulty in identifying one-egg human twins as undeniably separate personalities for two excellent reasons: first, physical separation is the essence of our vernacular definition of individuality (see following essay); second, human personalities are so subtly and pervasively shaped by complex environments of life (whatever the quirky similarities between one-egg twins reared apart) that each person follows a unique path.
With vastly greater rarity, the dividing cells of a fertilized egg begin to separate into two groups, but do not complete the process—and conjoined (or Siamese) twins develop. Conjoined twins span the entire conceivable range from a single individual bearing a few rudimentary parts of an imperfect twin, to superficially joined, complete individuals like Chang and Eng. Ritta and Christina fall squarely in the middle of this continuum. With our modern knowledge of their biological formation, I fear that we must reject Serres’s solution, and admit instead that his dilemma cannot be answered.
We inhabit a complex world. Some boundaries are sharp and permit clean and definite distinctions. But nature also includes continua that cannot be neatly parceled into two piles of unambiguous yeses and noes. Biologists have rejected, as fatally flawed in principle, all attempts by anti-abortionists to define an unambiguous “beginning of life,” because we know so well that the sequence from ovulation or spermatogenesis to birth is an unbreakable continuum—and surely no one will define masturbation as murder. Our congressmen may create a legal fiction for statutory effect but they may not seek support from biology. Ritta and Christina lay in the middle of another unbreakable continuum. They are in part two and in part one. And this, I am sorry to say, is the biological nonanswer to the question of the centuries.
If this argument leaves you with an empty feeling after so much verbiage, I can only reply with the paradoxical phrase that is, so often, the most liberating response to an old mystery: The question has no answer because you asked the wrong question. The old question of individuality in Siamese twinning rests upon the assumption that objects can be pigeonholed into discrete categories. If we recognize that our world is full of irreducible continua, we will not be troubled by the intermediate status of Ritta and Christina.
Dante punished schismatics by dismembering them in hell to exact a physical punishment worthy of their ideological crime: “Lo, how is Mohammed mangled…. Whom here thou seest, while they lived, did sow scandal and schism, and therefore thus are rent.”
Let us value connections. As Dante analogized physical with ideological separation, perhaps we can learn from the indissoluble union of Ritta and Christina that our intellectual world revels in continuity as well.
ABSTINENCE HAS
its virtuous side, but enough is enough. I have always felt especially sorry for poor Mabel, betrothed to Frederic the pirate apprentice. On the very threshold of married happiness, she discovers that she must wait another sixty-three years to claim her beloved at age eighty-four—and, as could only happen in Gilbert and Sullivan, she actually promises to wait.
The Pirate King and Ruth, Frederic’s old nursemaid and jilted paramour, present the reason for this extraordinary delay. Frederic, wrongfully apprenticed to the pirate band, has reached his twenty-first year and longs for freedom, respectability, and Mabel. But he was formally bound until his twenty-first
birthday
, and he was born on February 29. “You are a little boy of five,” the Pirate King informs him with glee and expectation of prolonged service. The three principals of the
Pirates of Penzance
then analyze the complexities of this predicament in the famous paradox song:
How quaint the ways of paradox
At common sense she gaily mocks.
The classic paradox presents us with two contradictory interpretations, each quite correct in its own context. Consider our western prototypes, the so-called paradoxes of Zeno: The arrow that can never reach its destination because, at any instant, it must occupy a fixed position; and Achilles who will never catch the tortoise because he must first traverse half the remaining distance, and any gap, no matter how small, can still be halved. We delight in paradox because it appeals to both the sublime and whimsical aspects of our psyche. We laugh with Frederic, but also feel that something deep about the nature of logic and life lies hidden in Zeno’s conundrums.
Biology too has its classical paradox. It flared as a major issue in the nineteenth century, probably because scientists then felt that they might find a resolution. All the best naturalists struggled in vain: Huxley and Agassiz lined up on opposite sides; Haeckel tried to mediate. The twentieth century has largely bypassed the conundrum, probably because we now realize that no simple answer exists. Yet, if our fascination with paradox be justified, the question can still enlighten us by virtue of its stubborn intractability.
Physalia
, the Portuguese man-of-war, embodies all this fuss. It is a siphonophore, a relative of corals and jellyfish. The old paradox addresses an issue that could not be more fundamental—the definition of an organism and the general question of boundaries in nature. Specifically: Are siphonophores organisms or colonies?
Siphonophores belong to the phylum Cnidaria (or Coelenterata). Two aspects of cnidarian biology set the context of our paradox. First, many cnidarians live as colonies of connected individuals—our massive coral reefs are gigantic congeries made of many million tiny, conjoined polyps. Second, the cnidarian life cycle features a so-called alternation of generations. The sessile polyp, a fixed cylinder with a fringe of tentacles, reproduces asexually and generates by budding the free-swimming medusa, or “jellyfish.” The medusa produces sexual cells that unite and grow into a polyp. And so it goes.
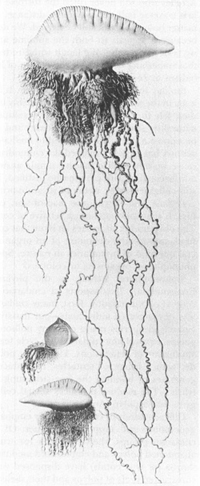
A Portuguese man-of-war. The creature is a colony, not a single organism. The float is a medusa person, and each “tentacle” is a polyp person.
FROM LOUIS AGASSIZ’S MONOGRAPH
(1862),
REPRINTED FROM NATURAL HISTORY
.

T.H. Huxley’s 1849 illustration of the Portuguese man-of-war. He regarded this creature as an individual, not a colony.
Different kinds of cnidarians may emphasize one of these generations and suppress the other. Of the three major cnidarian groups, the Scyphozoa (or true jellyfish) have abandoned polyps and emphasized medusae, while the Anthozoa (or true corals) have dispensed with medusae and constructed reefs of polyps and their skeletons. In the third group, the Hydrozoa, many members retain the full cycle, with prominent polyp and medusa. Siphonophores are hydrozoans. The technical literature, not usually noted either for charm or directness, has transcended its usual limitatons in this case: amidst a forest of formidable jargon for other parts of cnidarian anatomy, it refers to the polyp and medusa stages of a single life cycle as “persons.”
The Portuguese man-of-war, with its float above and tentacles below, looks superficially like a jellyfish (that is, a single medusa). When studied more carefully, we find that this floating weapon is a colony of many persons, both polyps and medusae. The pneumatophore, or float, is probably a greatly modified medusa (though some scientists think that it may be an even more altered polyp). The “tentacles,” though varied and specialized for different roles of capturing food, digestion, and reproduction, are not simple parts of a jellyfish but modified polyps—that is, each tentacle arises as a discrete person. (Another common siphonophore,
Velella
, literally the “little sail” but popularly given the lovely name of “by-the-wind sailor,” provokes even more confusion. Its persons are few enough and so well coordinated that the colony looks like a simple float surrounded by tentacles—in other words, like a simple jellyfish. But the float is a medusa person and each tentacle a polyp person.)
*
If this degree of division of labor among persons impresses you, nature has much more to offer.
Physalia
and
Velella
are simple siphonophores, with relatively few types of modified persons. The more complex siphonophores are, by far, nature’s most integrated colonies. Their parts are so differentiated and specialized, so subordinate to the entire colony, that they function more as organs of a body than persons of a colony.
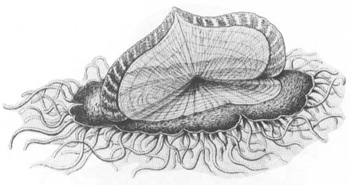
Velella
, the “by-the-wind-sailor,” is a colony—the float is a medusa person, each “tentacle” is a polyp person.
FROM E. HAECKEL’S
Challenger
MONOGRAPH
(1888).
REPRINTED FROM NATURAL HISTORY
.
Most siphonophores are small, transparent creatures of the open sea. They float at the surface among the plankton or swim actively, usually at shallow depths. As carnivores, they capture small planktonic animals in their net of tentacles. Larger siphonophores,
Physalia
among them, can ensnare and devour fish of substantial size; as many of us know to our sorrow, they can also inflict painful stings upon human bathers.
Complex siphonophores include an imposing array of well-differentiated structures. Their bodies may be roughly divided into two parts: an upper set of bulbs and pumps for locomotion and a lower set of tubes and filaments for feeding and reproduction. Each part contains a series of different polyps and medusae.
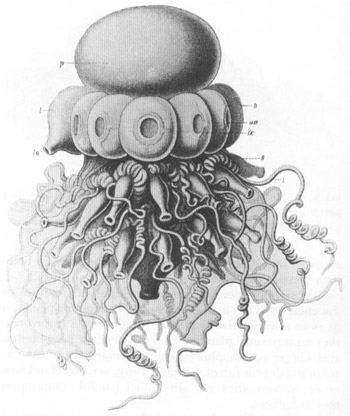
A relatively “simple” representative of the complex siphonophores, just for starters. Only four basic types of individuals are shown—two upper kinds of medusa persons (the pneumatophore, or float, labeled p; and a row of nectophores, or swimming bells, labeled n); and two lower kinds of polyp persons (the feeding siphons, labeled s; and the long sensory tentacles, labeled t).
FROM HAECKEL’S
Challenger
MONOGRAPH
, 1888.
Consider first the range of forms and activities assumed by polyp persons. We find three basic types and a myriad of modifications. The feeding organs, or siphons (hence the group’s name—
siphonophore
means “siphon bearer”), are tubular structures each with a stomach and trumpet-shaped mouth, usually hanging in profusion below the floats and swimming persons. The siphons are minimally modified polyp persons, and we can easily comprehend their origin as complete organisms. All other types of polyps (and most medusae) are more highly altered and specialized, and therefore more difficult to link with their original personality. A second order of polyp persons, the so-called dactylozooids (“finger,” or touching, animals), capture and transport food to the siphons. Dactylozooids include the extended thin tentacles, sometimes more than fifty feet long in
Physalia
, that carry the painful nematocysts, or stinging cells, and form a transparent web to ensnare prey. They have retained neither mouth nor digestive apparatus and might easily be taken for parts rather than persons if we could not trace their origin as discrete buds in growth.
These capturing parts often display a remarkable complexity of form and function. The stinging cells may be concentrated into swellings, or “batteries,” sometimes protected by a hood. In
Stephanophyes
, each battery ends in a delicate terminal filament and contains about 1,700 stinging cells of four different types. The terminal filament lassoes the prey and discharges its few stinging cells. If these cells fail to dispatch the victim, the filament contracts and carries the prey to the far end of the battery itself, where another volley of larger stinging cells transfixes the victim. If the prey continues to struggle, another contraction moves it up the battery to the near end, where the largest and most powerful stinging cells finally end the torment before passing the vanquished prey along to the siphon for ingestion.
Jennifer E. Purcell (see bibliography) has recently presented further evidence that feeding and capturing persons do not form a simple passive network, like the web of a spider, but play an active role in obtaining food. She found that the stinging cell batteries of two species function as lures by resembling, in both form and motion, small zoo-plankton that serve as prey for animals eaten by siphonophores. The batteries of
Agalma okeni
look like a copepod with two long antennae; each contracts independently at varying intervals of five to thirty seconds, creating a web of motion that simulates the darting and swimming of a copepod school (or whatever you call an aggregation of these tiny planktonic arthropods). To seal the story, Purcell opened the stomachs of
Agalma
and found the remains of three creatures, all predators of copepods. The batteries of another species,
Athorybia rosacea
, resemble the planktonic larvae of fish. They also contract rapidly, mimicking the swimming and feeding motions of their models.
Gonozooids, the third category of polyp persons, are reproductive structures. They are usually short, simple tubes, without mouth or motion. But they bud off the medusa persons, which then make reproductive cells to produce the next generation of siphonophores.