Eight Little Piggies (45 page)
Read Eight Little Piggies Online
Authors: Stephen Jay Gould

Patten’s iconography (see figure) portrays this claim for linear progress as a single grand highway. This figure is unconventional in its dubious attempt to compress three dimensions into two. The vertical axis represents “progress in organization, brain size, parental care, and adaptability,” rather than the usual geological time. Instead, time runs along the horizontal axis, symmetrically in both directions, from the origin of life at the center, to younger and younger strata both right and left. But time correlates with progress (the chart would be unintelligible otherwise)—so farther from the beginning point (bottom, center) means younger and better. Thus, Patten draws four bubbles, outlined in black and shaped like light bulbs, centered on the starting point. Each bubble adds geologically younger and biologically more advanced forms. The four bubbles represent, in order, Pre-cambrian, Paleozoic, Mesozoic, and Cenozoic eras. The third dimension of organic form can’t be plotted into this scheme, but Patten fudges and simply draws the conventional spindle for each group, radiating from the interior of a bubble toward its edge.
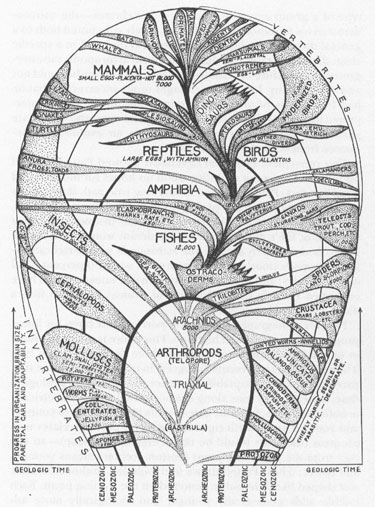
Patten’s progressivist tree of animal evolution. Note how arthropods progressed directly to vertebrates through ostracoderm intermediates.
William Patten’s picture from his
Evolution.
Plate 1, Dartmouth Press, 1925
.
Note the major feature of this iconography—and the obvious rationale behind Patten’s chosen form: The Great Highway of Animal Evolution ascends right through all the bubbles in the center of the chart. It rises from arthropods in the first bubble; to ostracoderms (the crucial link, including
Cephalaspis
), fishes, and amphibians in the second; to dinosaurs and early mammals in the third bubble; and finally to man, flanked by primates and hoofed mammals, in the fourth bubble. The great highway does become dangerously constricted now and then, but it always perseveres and always moves like the motto of New York State and the skyscrapers of its metropolis—Excelsior, “ever upward.”
How shall we compare Patten’s iconography with Agassiz’s earlier chart? Is it truer? Better? Does it represent a simple accretion of knowledge—scientific progress—in the intervening eighty years? I suppose so, in some sense. Patten does connect the spindles to acknowledge the discovery of evolution. He also recognizes the coherence of ostracoderms as an ancestral group of vertebrates, whereas Agassiz had regarded
Cephalaspis
alone as a confusing oddity. Yet in other ways, Patten’s chart has lost accuracy. Agassiz correctly classified
Cephalaspis
as a fish, whereas Patten, impelled by his theory of necessary progress, managed to reconstruct the ostracoderms as part arthropod.
But more fundamental than these backings-and-forthings must be the basic incommensurability of these two charts. They are not linearly related by progress or regress in knowledge. In fact, you cannot transform one into the other. They represent two incompatible worldviews, not a filling-in of new facts on the objective background of history. Each scheme for relationships among organisms depicts a personal theory, not a hatrack stamped out of universal logic. And the main difference isn’t even the watershed that we usually identify as the chief contrast between Agassiz’s and Patten’s worlds—the intervening discovery of evolution. Agassiz’s topology is easily converted to an evolutionary scheme by connecting the spindles. But Agassiz’s theory cannot be transformed into Patten’s worldview, for Agassiz based his vision on differentiation (radiation of numerous lineages from common points of origin), while Patten embraced linear progress. You can’t turn a hand into an upraised forearm.
When Stensiö resolved the debate about
Cephalaspis
by proving both its jawlessness and its relationship to modern lampreys and hagfishes, he summarized his discoveries in a third iconography. This more modest chart (see figure) does not show all life (like Patten’s) or even all fishes (like Agassiz’s); it portrays jawless fishes only—both the ostracoderms and their modern descendants. It argues that a root stock of ostracoderms split into two basic groups: the first containing two groups of fossils (including
Cephalaspis
among the Osteostraci) and the modern lampreys (Petromyzontia); the second including two other fossil groups and the modern hagfishes (Myxinoidea).
Again, Stensiö’s iconography does not emerge by accretion of information onto Patten’s version. It represents yet another worldview, incommensurate with Patten’s and therefore not derivable by any transformation from Patten’s grand highway. Stensiö’s organizing scheme is diversification, not progress. His iconography is closer to Agassiz’s preevolutionary version than to Patten’s supposed improvement. Connect Agassiz’s spindles and you obtain something more like Stensiö’s evolutionary branching than Patten’s linear progress.
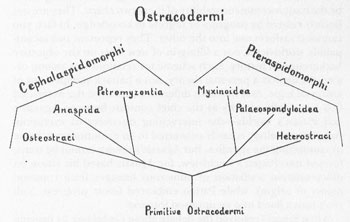
Stensiö’s evolutionary tree of ostracoderm fishes.
Erik Stensiö’s sketch from his monograph
Downtonian and Devonian Vertebrates of Spitsbergen
Pt., 1, 279, 1927
.
But Stensiö is not simply Agassiz after a game of connect the dots. We still note a basic incommensurability. On Agassiz’s chart, all subsidiary groups radiate from a central axis within each of the four divisions. No geologically younger group splits from a previous side branch; all point toward the central stem. This choice records Agassiz’s belief that creation occurs on an embryological plan. Mammals did not evolve directly from reptiles (themselves a side branch of the vertebrate trunk). Mammals were created after the death of ruling reptiles—as a more highly differentiated incarnation of the vertebrate ideal. The central stem of each group is, for Agassiz, the archetype that must serve as a model for each new and independent side branch of created complexity. But Stensiö’s iconography is fully evolutionary, with hierarchical diversification. Stems produce side branches, and side branches then bud off other twigs. Agassiz’s iconography is like a human hand; on Stensiö’s chart, fingers generate fingerlets, which generate fingerletchens, which generate…
These three successive iconographies lead me to conclude that scientific change cannot, in this case at least, be viewed as a simple accumulation of knowledge within the unchanging framework of a universal, objective method. We did learn more and more about
Cephalaspis
in particular and about the evolution of vertebrates in general. But the theories of Agassiz, Patten, and Stensiö are three incompatible worldviews—three visions imposed upon our greatly imperfect knowledge—not a progressive fleshing out of nature’s bare bones. And ironically, for those wedded to linear progress in ideas (if not in life’s history), Stensiö’s “final” version shares more of its iconography with Agassiz’s creationist vision of differentiation than with Patten’s evolutionary dream of linear advance.
Nature does not tolerate chimeras among its more complex animals. You cannot put a man’s head and chest upon a horse’s torso, and you certainly can’t meld an arthropod’s head with a vertebrate’s trunk (as Patten proposed in arguing that
Cephalaspis
had the jaw of a sea scorpion). Organic designs have integrity as working wholes constructed from coherent patterns of development. Nature is not an interior decorator or a postmodern builder recycling the entire history of architecture in an eclectic display of façades.
I believe that ideas have a similar integrity. Great thinkers build their edifices with subtle consistency. We do our intellectual forebears an enormous disservice when we dismember their visions and scan their systems in order to extract a few disembodied “gems”—thoughts or claims still accepted as true. These disarticulated pieces then become the entire legacy of our ancestors, and we lose the beauty and coherence of older systems that might enlighten us by their unfamiliarity—and their consequent challenge—in our fallible (and complacent) modern world.
This integrity of systems also precludes smooth transitions in intellectual life. Some systems will not transform smoothly into others, and discontinuous change must occur from time to time in the history of ideas, of human social institutions, and in the form of organisms. D’Arcy Thompson, great morphologist and admirer of Goethe (see Essay 10), made his idiosyncratic argument for rapid transitions in his classic treatise
On Growth and Form
(1917). He probably exaggerated the case for organisms with his geometric analogy, but I recommend his words as a stimulating, if overstated, model for the history of great ideas:
An algebraic curve has its fundamental formula, which defines the family to which it belongs…. We never think of “transforming” a helicoid into an ellipsoid, or a circle into a frequency-curve. So it is with the forms of animals. We cannot transform an invertebrate into a vertebrate, nor a coelenterate into a worm, by any simple and legitimate deformation…. Nature proceeds from one type to another…and these types vary according to their own parameters, and are defined by physico-mathematical conditions of possibility…. To seek for stepping-stones across the gaps between is to seek in vain, forever.
FRANCIS GALTON
, Darwin’s cousin and England’s most eccentric scientific genius, took all knowledge (and much speculation) for his province. Many of his studies were cranky or dubious. He performed a statistical test on the efficacy of prayer and, as inventor of the word
eugenics
, led a movement for selective marriage among the élite. But he could also be outstandingly right in his originality, as in his successful campaign for using fingerprints to identify criminals.
In his autobiography, Galton tells a story of Herbert Spencer’s visit to his fingerprint lab. Galton took Spencer’s prints and “spoke of the failure to discover the origin of these patterns, and how the fingers of unborn children had been dissected to ascertain their earliest stages.” Spencer, quick to offer certain opinions about almost anything, told Galton that he had been working the wrong way round.
Spencer remarked…that I ought to consider the purpose the ridges had to fulfil, and to work backwards. Here, he said, it was obvious that the delicate mouths of the sudorific glands required the protection given to them by the ridges on either side of them, and there from he elaborated a consistent and ingenious hypothesis at great length. I replied that his arguments were beautiful and deserved to be true, but it happened that the mouths of the ducts did not run in the valleys between the crests, but along the crests of the ridges themselves.
Galton then ends his anecdote by giving the original source for one of the top ten among scientific quotes. Spencer, dining with T. H. Huxley one night at the Athenaeum, stated that he had once written a tragedy. Huxley replied that he knew all about the work. Spencer rebutted Huxley, arguing that he had never mentioned it to anyone. But Huxley insisted that he knew anyway and identified Spencer’s debacle—“a beautiful theory, killed by a nasty, ugly little fact.”
Some beliefs may be subject to such instant, brutal, and unambiguous rejection. For example, no left-coiling periwinkle has ever been found among millions of snails examined. If I happen to find one during my walk on Nobska beach tomorrow morning, a century of well-nurtured negative evidence will collapse in an instant.
This Huxleyan vision of clean refutation buttresses one of our worst stereotypes about science. We tend to view science as a truth-seeking machine, driven by two forces that winnow error: the new discovery and the crucial experiment—prime generators of those nasty, ugly little facts. Science does, of course, seek truth, and even succeeds reasonably often, so far as we can tell. But science, like all of life, is filled with rich and complex ambiguity. The path to truth is rarely straight, marked by a gate of entry that sorts applicants by such relatively simple criteria as age and height. (When I was a kid, you could get into Yankee Stadium for half price if your head didn’t reach a line prominently drawn on the entrance gate about four feet above the ground. You could scrunch down, but they checked. One nasty, ugly day, I started to pay full price, and that was that.)
Little facts rarely undo big theories all by themselves—the myth of David and Goliath notwithstanding. Such facts can refute little, highly specific theories, like my conjecture about lefty periwinkles, but they rarely slay grand and comprehensive views of nature. No single, pristine fact taught us that the earth revolves around the sun or that evolution produced the similarities among organisms. Overarching theories are much bigger than single facts, just as the army of Grenada really didn’t have much chance against the combined forces of the United States.
Instead, little facts are assimilated into large theories. They may reside there uncomfortably, bothering the honorable proponents. Large numbers of little facts may eventually combine with other social and intellectual forces to topple a grand theory. The history of ideas is a play of complex human passions interacting with an external reality only slightly less intricate. We debase the richness of both nature and our own minds if we view the great pageant of our intellectual history as a compendium of new information leading from primal superstition to final exactitude. We know that the sun is hub to our little corner of the universe, and that ties of genealogy connect all living things on our planet, because these theories assemble and explain so much otherwise disparate and unrelated information—not because Galileo trained his telescope on the moons of Jupiter or because Darwin took a ride on a Galápagos tortoise.
This essay tells the story of a pristine, unexpected little fact that should have mattered, but didn’t particularly. The fact was widely reported, discussed, and personally studied by the greatest naturalists of Europe, and then assimilated into each of several contradictory systems. Fifty years later, in 1865, a second discovery resolved the paradox generated by the first fact—and should have won, by Huxley’s principle, a big and important victory for Darwin and evolution. It was welcomed, to be sure, but largely ignored. One foot soldier could not decide a battle waged on so many fronts.
Trigonia
is a distinctive clam, thick shelled and triangular in shape. It flourished with dinosaurs during the Mesozoic era and then became extinct in the same debacle that wiped out the ruling reptiles—one of the five greatest mass dyings in our geologic record. No trigonian had ever been found in the overlying Cenozoic strata—the entire age of mammals (about sixty million years, as we now know).
Trigonia
had therefore become a valued “guide fossil” when you found one, you knew you had rocks of the earth’s middle age. Everyone (who was anybody) understood this useful restriction in time.
Then, the nasty, ugly little—and quite undeniable—fact: In 1802, P. Péron, a French naturalist, found the shell of a living trigonian washed up on the beaches of southern Australia. Twenty-five years later, and following several failures, J. Quoy and J. Gaimard, naturalists aboard the
Astrolabe
, finally found a live trigonian. They had dredged for several days with definite purpose, but without success. Becalmed one night in Bass Strait and with little else to do, they tried again and brought up their single prize, a molluscan life soon snuffed and preserved in the medium of the collector’s trade—a bottle of alcohol. Quoy and Gaimard treasured their booty and wrote later:
We were so anxious to bring back this shell with its animal that when we were, for three days, stranded on the reefs of Tonga-Tabu, it was the only object that we took from our collection. Doesn’t this recall the ardent shell collector who, during seven years’ war, carried constantly in his pocket an extraordinary
Phasianella
, which he had bought for twenty-five louis?
A simple story. A fact and a puzzle.
Trigonia
had not disappeared in the great Cretaceous debacle, but lived still in Australia. Yet no fossil trigonians had been found in all the strata in between—throughout the long and well-recorded history of the age of mammals (now called the Cenozoic era). Where were they? Had they ever existed? Could such a distinctive animal die and be reborn (or re-created) later? The “Cenozoic gap” became as puzzling and portentous as the one later associated with Mr. Nixon and Ms. Woods.
Trigonia
occupies a specially interesting place in the history of biology because its unexpected fact and consequent puzzle arose and prevailed at such an important time—at the dawn and through the greatest conceptual transition ever experienced by the profession: from creationist to evolutionary views of life.
Trigonia
also (or rather therefore) attracted the attention and commentary of most leaders in nineteenth-century natural history. J. B. Lamarck, most famous of pre-Darwinian evolutionists, formally described the first living trigonian. Darwin himself thought and commented about
Trigonia
for thirty years. Louis Agassiz, most able and cogent of Darwin’s opponents, wrote the major technical monograph of his generation on the genus
Trigonia
.
The lesson of the living
Trigonia
can be distilled in a sentence: Everyone made the best of it, incorporating favorable aspects of this new fact into his system and either ignoring or explaining away the difficulties.
Trigonia
became an illustration for everyone, not a crucial test of rival theories. Evolutionists celebrated the differences in form and distribution between ancient and modern trigonians—and ignored the Cenozoic gap. Creationists highlighted the gap and made light of the differences.
Today, we remember Lamarck best as the author of a rejected evolutionary theory based on the inheritance of acquired characters (quite an unfair designation since so-called Lamarckian inheritance represents a minor part of Lamarck’s own system). But his day-to-day work in post-revolutionary France focused on the description of living and fossil invertebrates in his role as curator at the
Muséum d’Histoire Naturelle
in Paris. He therefore received Péron’s precious shell for formal description, and he named it
Trigonia margaritacea
in 1804 (Lamarck didn’t know about modern cocktails; margarita is a Latin pearl, and the interior of a trigonian shell shines with a beautiful pearly luster). But since 1804 lay squarely between Lamarck’s initial (1802) and definitive (1809) statement of his evolutionary theory, he also used his short paper on
Trigonia
to sharpen and defend his developing transmutationist views.
Most fossil trigonians are ornamented with concentric ridges at their anterior ends (enclosing the mouth and digestive apparatus) and radial ribs on the rear flank. A single strong rib usually separates these two areas. But all modern trigonians cover their shells entirely with radial ribs (although the embryonic shell still bears traces of the ancestral concentrics). Lamarck seized upon these differences to claim that changing environments had pressed their influence upon the shell. The shell had then altered in response and the animal within passed the favorable change to future generations by “Lamarckian” inheritance.

Lamarck’s original figure of the shell of living trigonian clams.

Fossil trigonians (left) have both concentric and radial ribs. Living trigonians (right) have only radial ribs.
They have undergone changes under the influence of circumstances that act upon them and that have themselves changed; so that fossil remains…of the greatest antiquity may display several differences from animals of the same type living now but nevertheless derived from them.
(But Lamarck had only demonstrated that the fossils looked different from the modern shells. Any theory could account for this basic datum in the absence of further information—evolution by use and disuse, by natural selection, or even re-creation by God for that matter.)
Lamarck then proceeded to extract more from modern trigonians to buttress other pet themes. He was, for example, a partisan at the wrong end of a great debate resolved a decade later to his disadvantage by Cuvier—does extinction occur in nature? Human rapacity, Lamarck believed, might exterminate some conspicuous beasts, but the ways of nature do not include termination without descent (Lamarck, as a transmutationist, obviously accepted the pseudoextinction that occurs when one form evolves into another). Lamarck gave the old arguments against extinction a novel twist by embedding his justification within his newfangled evolutionary views. How can extinction occur if all organisms respond creatively to changing environments and pass their favorable responses to future generations in the form of altered inheritance?
Yet Lamarck’s conviction was sorely challenged by burgeoning data in his own field of marine invertebrate paleontology. So many kinds of fossils are confined to rocks of early periods. Where are their descendants today? Lamarck offered the only plausible argument in a world with few remaining terrae incognitae—they live still in the unexplored depths of the sea. Since Lamarck reveals his own discomfort with such an ad hoc solution by repeating it too often and too zealously—recall Shakespeare’s “the lady doth protest too much, me-thinks”—we may take as genuine his delight in
Trigonia
as a real case for a generalization devoutly to be wished: “Small species, especially those that dwell in the depths of the sea, have the means to escape man; truly among these we do not find any that are really extinct.” Lamarck then ends his paper by predicting that a large suite of creatures apparently extinct will soon be found at oceanic depths. We are still waiting.